Inbreeding Depression Explained
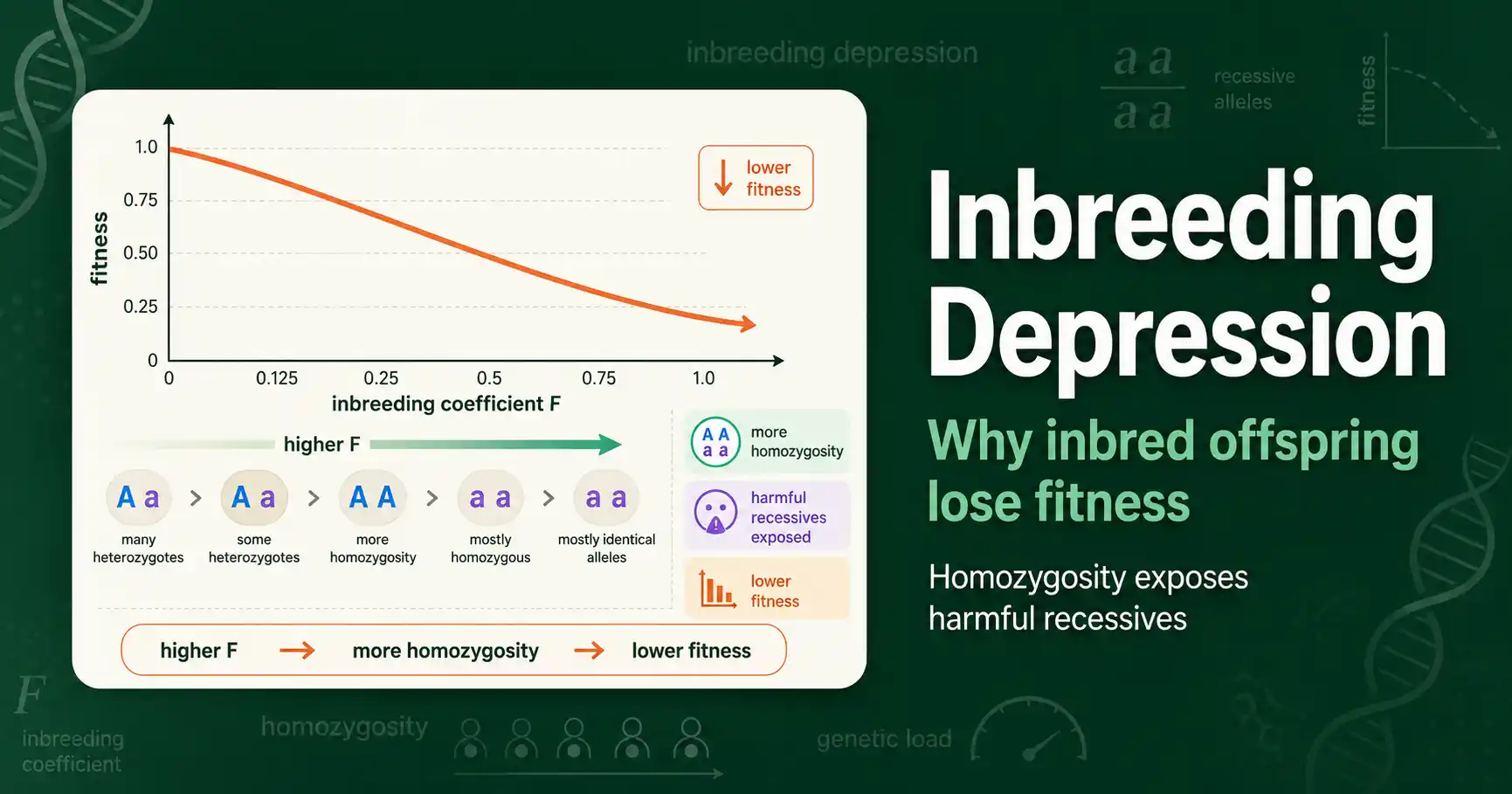
Inbreeding depression is the reduction in fitness, survival, and fertility seen in the offspring of related parents. The more inbred an individual is, the lower its expected fitness. This is why inbreeding matters in the first place, and it shows up across plants, animals, and humans alike.
The cause is homozygosity. Inbreeding makes individuals more likely to carry two identical copies of a gene, and that exposes harmful recessive alleles that normally stay hidden. This guide explains the mechanism, the genetics behind it, how strong the effect is, and why it drives decisions in conservation and breeding. The single number that predicts it is the inbreeding coefficient, covered in our explainer on what the inbreeding coefficient is.
What Inbreeding Depression Is
Inbreeding depression is a decline in the average quality of inbred individuals compared to outbred ones. The traits it hits hardest are the ones tied to survival and reproduction: fertility, offspring survival, growth, disease resistance, and developmental stability.
The effect scales with the inbreeding coefficient F. An individual with a higher F suffers more, on average, because a higher F means more of its genome is homozygous. The child of full siblings, at an F of 0.25, faces a larger expected fitness cost than the child of first cousins, at an F of 0.0625. The relationship is roughly linear for mild inbreeding, so fitness tends to fall in proportion to F.
This is a population-level average, not a guarantee for any one individual. Some inbred individuals are perfectly healthy, because whether the harmful recessives happen to pair up is partly chance. But across many inbred individuals, the average fitness decline is consistent and measurable, which is what makes inbreeding depression a real and predictable phenomenon rather than just bad luck.
Inbreeding depression also hits some traits much harder than others. Traits closely tied to fitness, survival to adulthood, fertility, and the number of offspring produced, show the strongest depression, because they depend on many genes and are sensitive to any harmful recessives that surface. Traits less directly tied to fitness, like minor morphological measurements, show weaker effects. This pattern is itself evidence for the dominance mechanism: fitness traits accumulate the most deleterious mutations over evolutionary time, so they have the most hidden load to expose when inbreeding strikes. A breeder or biologist looking for inbreeding depression therefore watches reproduction and survival first, since that is where the cost shows up earliest and most clearly.
Why Homozygosity Lowers Fitness
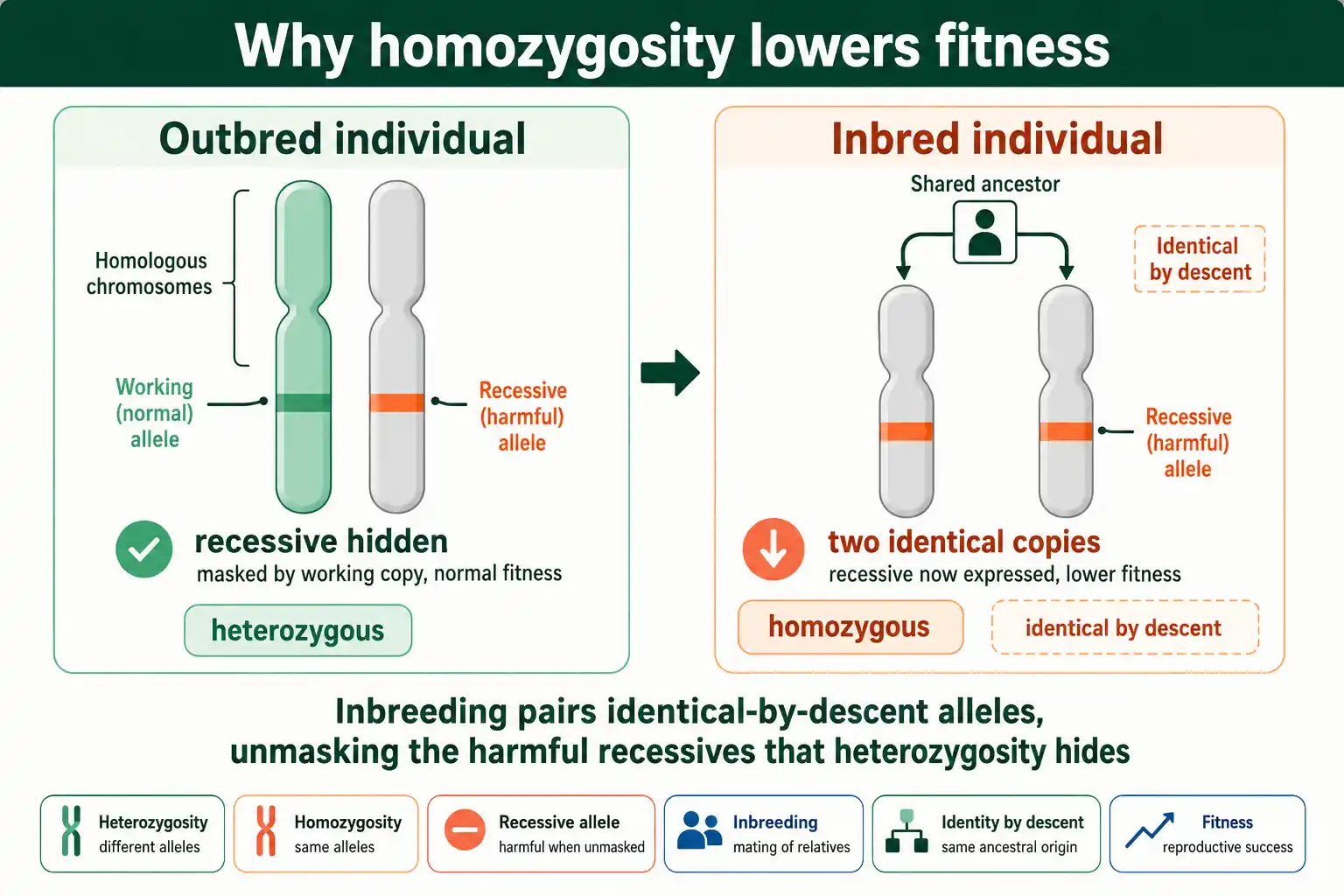
The root cause is simple: inbreeding raises homozygosity, and homozygosity exposes harmful alleles that heterozygosity hides. Two mechanisms explain how, and they have been debated for a century.
The first and main mechanism is the dominance hypothesis. Most harmful mutations are recessive or partly recessive, so they only damage fitness when an individual carries two copies. In an outbred individual, a single harmful recessive is masked by a working allele on the other chromosome. Inbreeding brings together two copies of the same ancestral allele, including harmful ones, so recessives that were hidden become expressed. The decline in fitness is the sum of all these newly exposed harmful recessives across the genome. As Deborah Charlesworth and John Willis established in their authoritative 2009 review in Nature Reviews Genetics, the dominance hypothesis, the unmasking of partially recessive deleterious mutations, is now accepted as the major cause of inbreeding depression.
The second mechanism is overdominance, where the heterozygote is fitter than either homozygote. Where overdominance operates, simply becoming homozygous lowers fitness even without harmful recessives, because the heterozygous advantage is lost. Overdominance contributes in some cases, but the long debate of the 1970s settled in favor of dominance as the primary driver, with overdominance playing a smaller role. To see the homozygosity side of this directly, our guide on heterozygosity covers how genetic diversity is measured and lost.
The Two Mechanisms Compared
The dominance and overdominance hypotheses make different predictions, which is how researchers tell them apart. The table lays out the contrast.
| Feature | Dominance hypothesis | Overdominance hypothesis |
|---|---|---|
| Cause of fitness loss | Exposure of harmful recessive alleles | Loss of heterozygote advantage |
| Allele effect | Deleterious alleles partly recessive | Heterozygote fitter than both homozygotes |
| Can it be purged? | Yes, selection can remove exposed recessives | No, fitness keeps falling with homozygosity |
| Current standing | Accepted as the major cause | Minor contributor in most cases |
The purging difference is the most useful one in practice. Under dominance, sustained inbreeding can actually expose harmful recessives to selection so effectively that they get removed from the population, a process called purging. Under overdominance, no such relief occurs, because the problem is homozygosity itself, not specific bad alleles. This is why purging behavior in real populations is taken as evidence for the dominance mechanism.
Charles Darwin Got There First
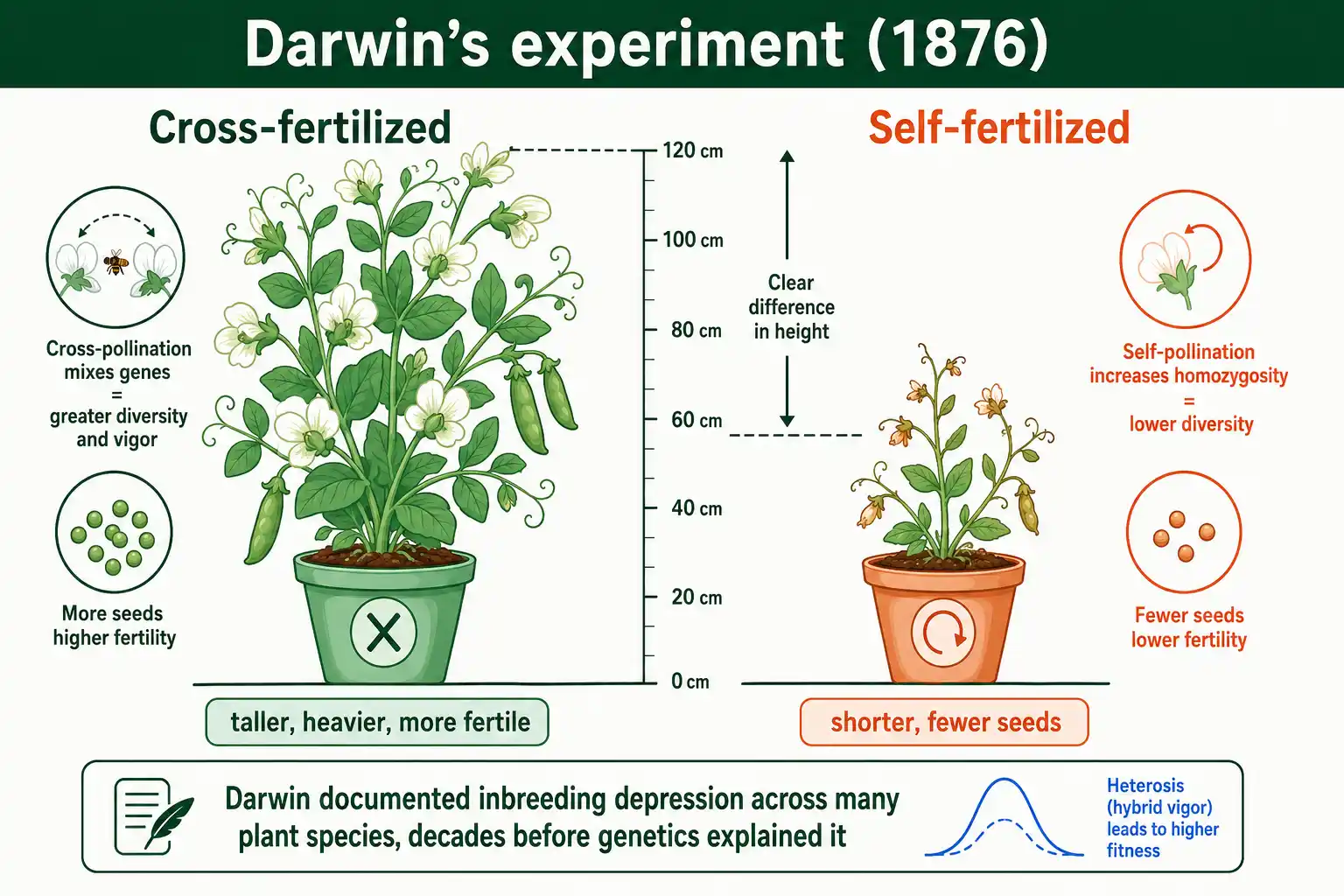
Inbreeding depression was documented long before genetics existed to explain it. Charles Darwin studied it systematically and published his findings in The Effects of Cross and Self Fertilisation in the Vegetable Kingdom in 1876.
Darwin ran experiments on plants, comparing the offspring of self-fertilized plants with those of cross-fertilized ones across many species. The cross-fertilized offspring were consistently taller, heavier, and more fertile. He had discovered inbreeding depression and its mirror image, the vigor of outcrossed offspring, without any knowledge of genes or alleles. His conclusion, that nature works against perpetual self-fertilization, anticipated the modern understanding by decades.
The genetics that explained Darwin's results came later, with the rediscovery of Mendel and the rise of population genetics in the early twentieth century. Sewall Wright's inbreeding coefficient gave the phenomenon a number, and the dominance and overdominance hypotheses gave it a mechanism. But the core observation, that inbred offspring are less fit than outbred ones, traces straight back to Darwin's greenhouse experiments. The fact that he saw it so clearly, in so many species, is part of why inbreeding depression is considered one of the most general patterns in biology.
How Strong Is the Effect
The strength of inbreeding depression varies by species and trait, but the effect is often large, especially for close inbreeding. Real figures put it in perspective.
In humans, the children of first cousins face a measurably higher risk of recessive genetic disorders and somewhat higher infant mortality than the children of unrelated parents, though the absolute increase from cousin-level inbreeding is modest. For first-degree inbreeding, between siblings or parent and child, the effects are severe, with high rates of serious genetic conditions and reduced survival. You can put a number on the underlying F for any relationship using an inbreeding calculator, which is the figure these risks scale with.
In other species the effect is often dramatic. Inbred plant lines show reduced height, seed production, and germination. Inbred animal populations show lower fertility, smaller litters, weaker immune function, and higher juvenile mortality. The reason inbreeding depression is so widespread is that every species carries a load of harmful recessive alleles, hidden in the heterozygous state, that inbreeding inevitably brings to light. This hidden burden is called the genetic load.
How Inbreeding Depression Is Measured
Researchers quantify inbreeding depression by comparing the fitness of inbred and outbred individuals, and the comparison yields a single summary number. The most common approach plots a fitness trait against F and measures the slope.
The steeper the decline of a trait as F rises, the stronger the inbreeding depression. A common summary, the inbreeding depression coefficient, expresses how much fitness drops at a given level of inbreeding, often the fitness of inbred offspring relative to outbred ones. A value near zero means little inbreeding depression; a large value means inbred individuals fare much worse.
Geneticists also use a measure called lethal equivalents, which estimates how many harmful alleles a population effectively carries, each of which would be lethal if made homozygous. A population with more lethal equivalents shows steeper inbreeding depression, because inbreeding exposes more of that hidden load. Human populations carry an estimated handful of lethal equivalents per person on average, which is why close inbreeding raises disease risk noticeably while distant relationships add little. These measures turn the abstract idea of a hidden genetic load into numbers that breeders and conservationists can compare across populations.
Evidence From Wild Populations
Inbreeding depression is not just a laboratory or pedigree phenomenon; it has been documented repeatedly in wild populations, which is some of the strongest evidence that it shapes real evolution.
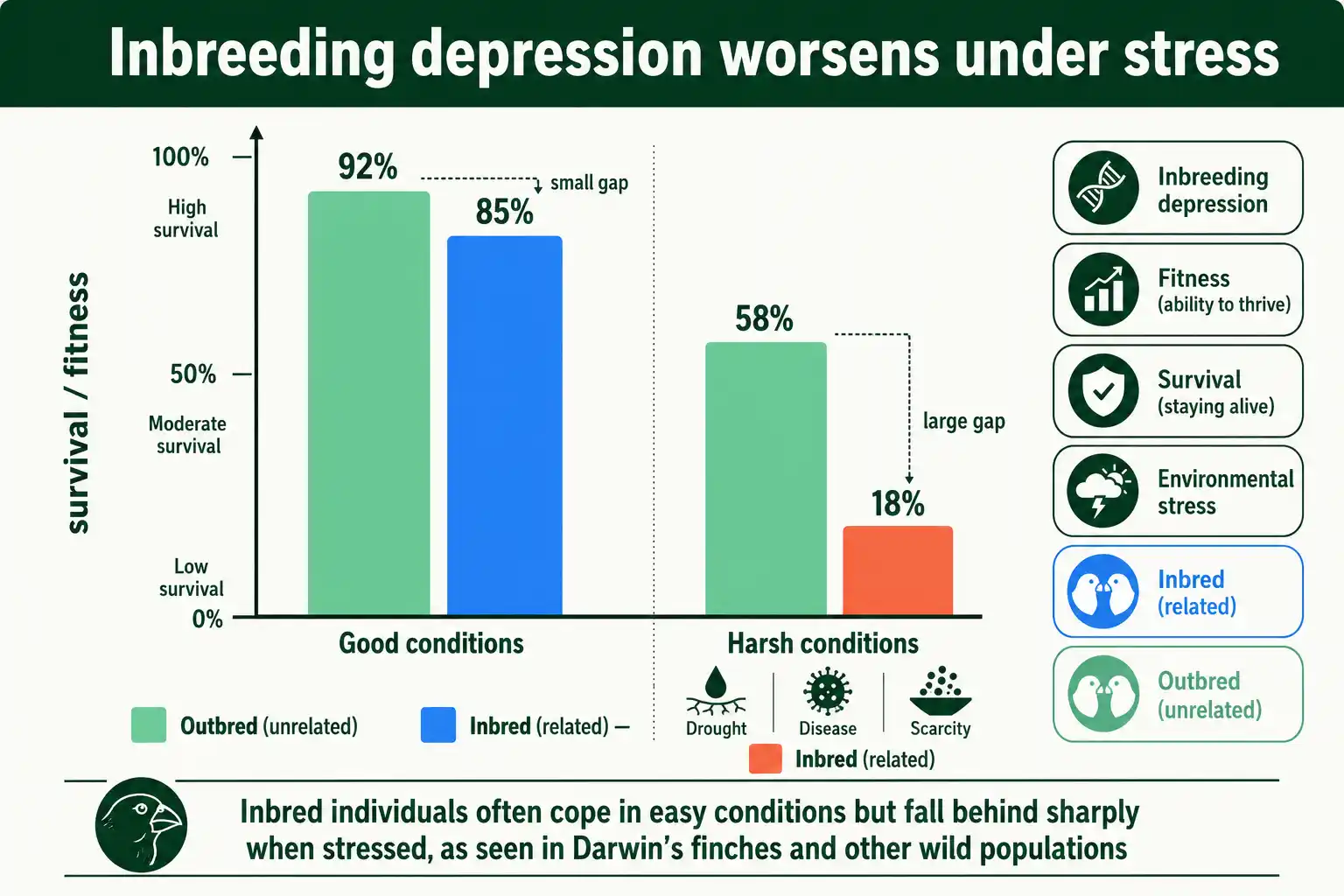
A celebrated example comes from the Darwin's finches of the Galapagos, where long-term field studies tracked individual birds across generations. More inbred finches showed lower survival and reduced reproductive success, and the effect was strongest in harsh years, revealing that inbreeding depression can intensify under environmental stress. The same stress-dependence appears in many species: inbred individuals often cope adequately in easy conditions but fall behind sharply when food is scarce or disease strikes.
Small and isolated populations provide stark cases. The Florida panther, reduced to a few dozen individuals, developed heart defects, poor sperm quality, and other signs of severe inbreeding depression before a genetic rescue restored diversity. Isolated populations of Scandinavian wolves, Swedish adders, and song sparrows on small islands have all shown measurable inbreeding depression in survival or reproduction. These cases matter because they confirm that inbreeding depression operates in nature, not only in controlled crosses, and that it is a genuine force pushing small populations toward decline.
Purging: Can Inbreeding Cure Itself?
A counterintuitive idea follows from the dominance mechanism: sustained inbreeding can sometimes reduce inbreeding depression over time, through purging. Understanding its limits matters for breeding and conservation.
Purging works like this. When inbreeding exposes harmful recessive alleles as homozygotes, natural selection can see them and remove them, because the individuals carrying them have lower fitness and leave fewer offspring. Over many generations of inbreeding, the most harmful recessives get weeded out, and the population's genetic load falls. This is why some highly self-fertilizing plants and long-inbred laboratory strains show less inbreeding depression than their history would suggest.
But purging is unreliable, and counting on it is risky. It works best against severe, highly recessive alleles, which selection removes efficiently. It works poorly against the many mildly harmful alleles that make up most of the genetic load, because selection against each is weak, and genetic drift in small populations can even fix harmful alleles instead of removing them. So purging can blunt inbreeding depression in some cases, but it rarely eliminates it, and in small populations the harmful effects of drift often outweigh any purging benefit. Conservation programs cannot assume a threatened population will simply purge its way to health.
Inbreeding Depression in Humans
Humans show inbreeding depression like any other species, and the medical-genetics view of it is the most relevant for many readers. The effect is real, well documented, and proportional to F, while being modest at the cousin level that most consanguinity questions involve.
The clearest signal is recessive genetic disease. When parents share ancestry, their child is more likely to inherit two copies of the same rare recessive allele, so consanguineous couples have a higher chance of a child with a recessive condition. The increase is steep for close relationships and mild for distant ones, tracking F directly. For first cousins, studies generally find the risk of a serious birth defect or genetic disorder is roughly a few percentage points higher than the population baseline, a real but limited increase. For first-degree relationships, the risk is far higher, which is why such unions are associated with severe outcomes.
Two points keep this in perspective. First, the baseline matters: every pregnancy carries some risk of a genetic condition, and cousin-level consanguinity adds a modest increment on top, rather than creating risk from nothing. Second, the specific recessive conditions that matter depend on what alleles a particular family carries, which is why population history and family history both feed into real risk. This is exactly the kind of assessment a genetic counselor performs, combining the calculated F with knowledge of the conditions that run in a family or population.
Carrier screening has changed the picture in recent years. Couples who know they are related can now be tested for the specific recessive conditions they might both carry, turning a general statistical risk into concrete, actionable information. This is why the modern medical approach pairs the inbreeding coefficient, which sets the overall scale of risk, with targeted genetic testing, which identifies the actual conditions in play.
Why It Matters for Conservation and Breeding
Inbreeding depression is the practical reason F is tracked everywhere ancestry is managed. Two fields treat it as a central concern.
In conservation biology, inbreeding depression is a major threat to small and endangered populations. As a population shrinks, individuals are forced to mate with relatives, F rises, and inbreeding depression lowers survival and fertility, which shrinks the population further. This feedback can spiral toward extinction, a process the conservation literature calls the extinction vortex. Inbreeding depression, working alongside genetic drift, is one of the engines of that spiral, which is why restoring genetic diversity is a conservation priority, explored further in our guide on genetic drift in conservation.
In animal breeding, inbreeding depression sets a limit on how much breeders can concentrate desirable bloodlines. Breeding closely related animals fixes good traits but also raises F, surfacing breed-specific genetic disorders and reducing fertility and vigor. This is why studbooks track inbreeding and why breeders introduce unrelated stock periodically. The vigor that appears when two inbred lines are crossed, the reverse of inbreeding depression, is called heterosis or hybrid vigor, and it is exploited deliberately in crops like hybrid maize to boost yield. Inbreeding depression and heterosis are two sides of the same coin: the first is the cost of homozygosity, the second the benefit of restoring heterozygosity.
The conservation response to inbreeding depression is genetic rescue: introducing unrelated individuals into a small, inbred population to bring in new alleles and restore heterozygosity. The Florida panther recovery is the textbook success, where adding a handful of pumas from Texas reversed the signs of inbreeding depression within a generation. Genetic rescue works precisely because it undoes the homozygosity that caused the problem, masking the harmful recessives once again and lifting fitness. It is the most direct counter to inbreeding depression available to managers of small populations.
One caution tempers this. Crossing very divergent populations can occasionally cause the opposite problem, outbreeding depression, where offspring are less fit because locally adapted gene combinations get broken up or because the populations were genetically incompatible. So conservationists weigh the near-certain benefit of relieving inbreeding depression against the smaller risk of outbreeding depression, usually favoring rescue when a population is severely inbred. The balance between too little gene flow, which leaves inbreeding depression unchecked, and too much, which can erode local adaptation, is a recurring theme in managing small populations.
The Cost of Homozygosity
Inbreeding depression is the fitness cost that homozygosity imposes, and it is the reason the inbreeding coefficient matters at all. The mechanism is the exposure of harmful recessive alleles, hidden in heterozygotes but unmasked when inbreeding pairs two identical copies, with overdominance playing a smaller supporting role. The effect scales with F, hits survival and reproduction hardest, and appears across plants, animals, and humans, exactly as Darwin first documented in 1876 and as Charlesworth and Willis later explained genetically.
The phenomenon shapes the real world. It threatens small populations through the extinction vortex, limits how far breeders can inbreed desirable lines, and underlies the hybrid vigor that powers modern crop yields. Purging can soften it in some long-inbred lineages, but it offers no guarantee, especially in the small populations where the danger is greatest. For anyone weighing the risks of consanguinity in their own family, a genetic counselor or physician can assess the specific recessive conditions that matter for their situation. To connect the fitness cost back to the number that predicts it, our guide on the inbreeding coefficient from a pedigree shows how to read F from a family tree.