What Disrupts Hardy-Weinberg Equilibrium?
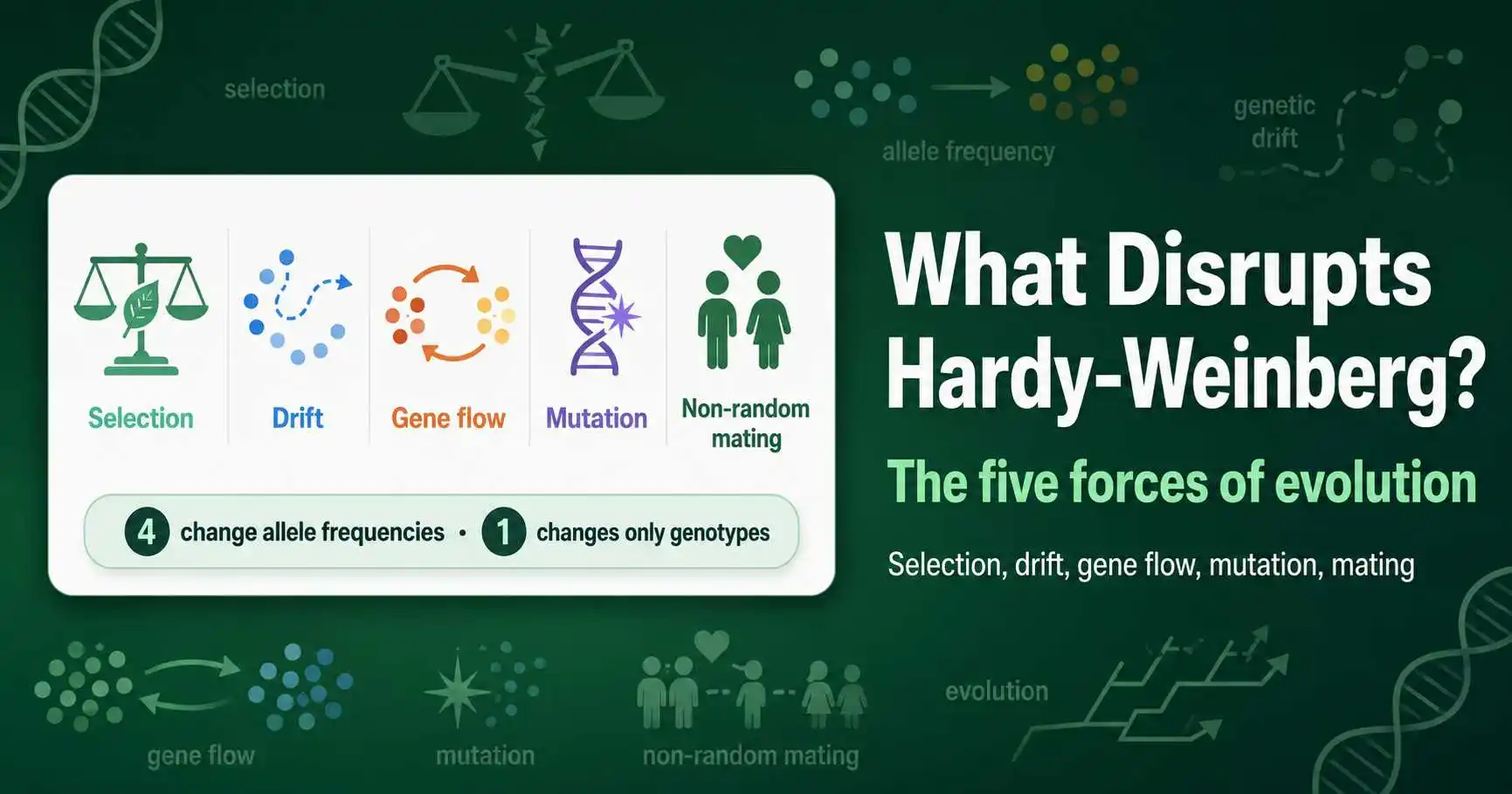
Five forces disrupt Hardy-Weinberg equilibrium: mutation, gene flow, genetic drift, non-random mating, and natural selection. The first four of these are genuine evolutionary forces that change allele frequencies, while non-random mating is a special case that changes only genotype frequencies. When any of these acts on a population, it actively pushes the population away from equilibrium, and that departure is exactly how scientists detect that evolution is occurring at a gene.
Each of the five conditions for Hardy-Weinberg equilibrium has a matching force that breaks it when the condition fails. This guide examines those forces as active processes, explaining how each one changes a population's genetics and illustrating each with a real-world example drawn from well-studied cases. It also draws out an important distinction that often gets lost: four of the forces alter allele frequencies, but one does not, which has real consequences for how it disrupts equilibrium. Equilibrium predictions can be modeled with a calculator, and understanding these forces explains why real populations deviate from those predictions.
Equilibrium as a Baseline, and What Breaks It
To understand what disrupts equilibrium, it helps to recall what equilibrium is. Hardy-Weinberg equilibrium is the state of a population that is not evolving, where allele and genotype frequencies stay constant across generations. It holds only under five idealized conditions, and it serves as a null model, the baseline of no change against which all real populations are compared.
The disrupting forces are simply the violations of those five conditions, viewed as active processes. Where the conditions are stated as absences, no mutation, no migration, large population, random mating, no selection, the forces are simply the corresponding presences: mutation, migration, drift, non-random mating, and selection. Each force, when present, undoes the stillness the model assumes, nudging or shoving the population away from its equilibrium frequencies. This is why studying the forces is the same as studying evolution itself, since these are precisely the mechanisms that drive genetic change over time, the processes that together account for how populations transform across generations.
A crucial framing makes the whole topic clearer. Four of the five forces, mutation, gene flow, genetic drift, and selection, change allele frequencies, which is the genetic definition of evolution. Non-random mating is the odd one out: it changes how alleles are packaged into genotypes but leaves the allele frequencies themselves untouched. So while all five disrupt the Hardy-Weinberg genotype predictions, only four of them cause evolution in the strict sense of changing allele frequencies. Keeping this distinction in view explains why non-random mating is sometimes described as not being a true evolutionary force at all. The conditions these forces violate are detailed in our guide to the assumptions of Hardy-Weinberg.
Natural Selection
Natural selection is the most famous disrupting force, and it changes allele frequencies whenever genotypes differ in their fitness, that is, their ability to survive and reproduce. Alleles that improve fitness become more common over generations, while alleles that reduce it become rarer, shifting the population away from equilibrium in a directed way.
The defining feature of selection is that it is not random. Unlike drift, which moves frequencies aimlessly, selection consistently pushes allele frequencies in the direction of higher fitness in the current environment. This makes it the engine of adaptive evolution, the process that fits organisms to their surroundings. Because the fitter genotypes leave more offspring, their alleles accumulate, and a trait that confers an advantage can spread through a population over relatively few generations when selection is strong.
The classic illustration is the peppered moth during the Industrial Revolution. Before heavy industry, light-colored moths were camouflaged against pale, lichen-covered tree bark, and dark moths were easily spotted and eaten by birds, so the light allele was common. As soot from factories darkened the bark, the situation reversed: dark moths were now better hidden and light moths stood out, so birds ate more light moths. The frequency of the dark allele rose sharply over a few decades, a clear, documented case of selection changing allele frequencies and driving a population out of Hardy-Weinberg equilibrium.
 When pollution controls later cleaned the air, the light form rebounded, showing selection tracking the environment in both directions.
When pollution controls later cleaned the air, the light form rebounded, showing selection tracking the environment in both directions.
When Selection Maintains Variation
Not all selection drives an allele toward fixation; some forms of selection actively preserve multiple alleles in a population, a pattern called balancing selection. This is worth a closer look because it produces a surprising result: a population under strong selection can still appear close to Hardy-Weinberg equilibrium.
The best-known example is the sickle cell allele in regions where malaria is common. Individuals with two copies of the sickle cell allele have sickle cell anemia, a serious condition, so that genotype is selected against. Individuals with two normal alleles are vulnerable to malaria. But heterozygotes, with one of each allele, are both protected against severe malaria and free of sickle cell anemia, giving them the highest fitness. This is called heterozygote advantage. Because the heterozygote is fittest, selection maintains both alleles in the population rather than eliminating either, keeping the sickle cell allele at a much higher frequency than its harmful homozygous effect would otherwise allow.
This case illustrates a subtle but important point about detecting selection. Balancing selection that favors heterozygotes can produce genotype frequencies that look close to the Hardy-Weinberg prediction, even though selection is clearly operating. This is why a population matching the equilibrium prediction does not absolutely prove that no force is acting. Selection is happening, yet the genotype distribution may not reveal it through a simple deviation. Heterozygote advantage is one of the clearest demonstrations that the relationship between equilibrium and evolution is not always straightforward, and it explains why some disease alleles persist at frequencies far higher than their harm alone would predict.
Genetic Drift
Genetic drift is the change in allele frequencies that happens purely by chance, and it is most powerful in small populations. Unlike selection, drift has no direction and no relationship to fitness; it is simply the random sampling error that occurs when a finite number of individuals pass on their alleles.
The mechanism is best understood as sampling. Each generation is a sample of alleles drawn from the previous one, and small samples can stray far from the true proportions by luck alone, just as a few coin flips can deviate widely from fifty-fifty. In a large population, these random deviations average out and frequencies stay stable. In a small population, chance can shift frequencies substantially each generation, and over time an allele may drift all the way to fixation, reaching 100 percent, or to loss, disappearing entirely. Drift reduces genetic variation and does so without regard to whether the surviving alleles are beneficial.
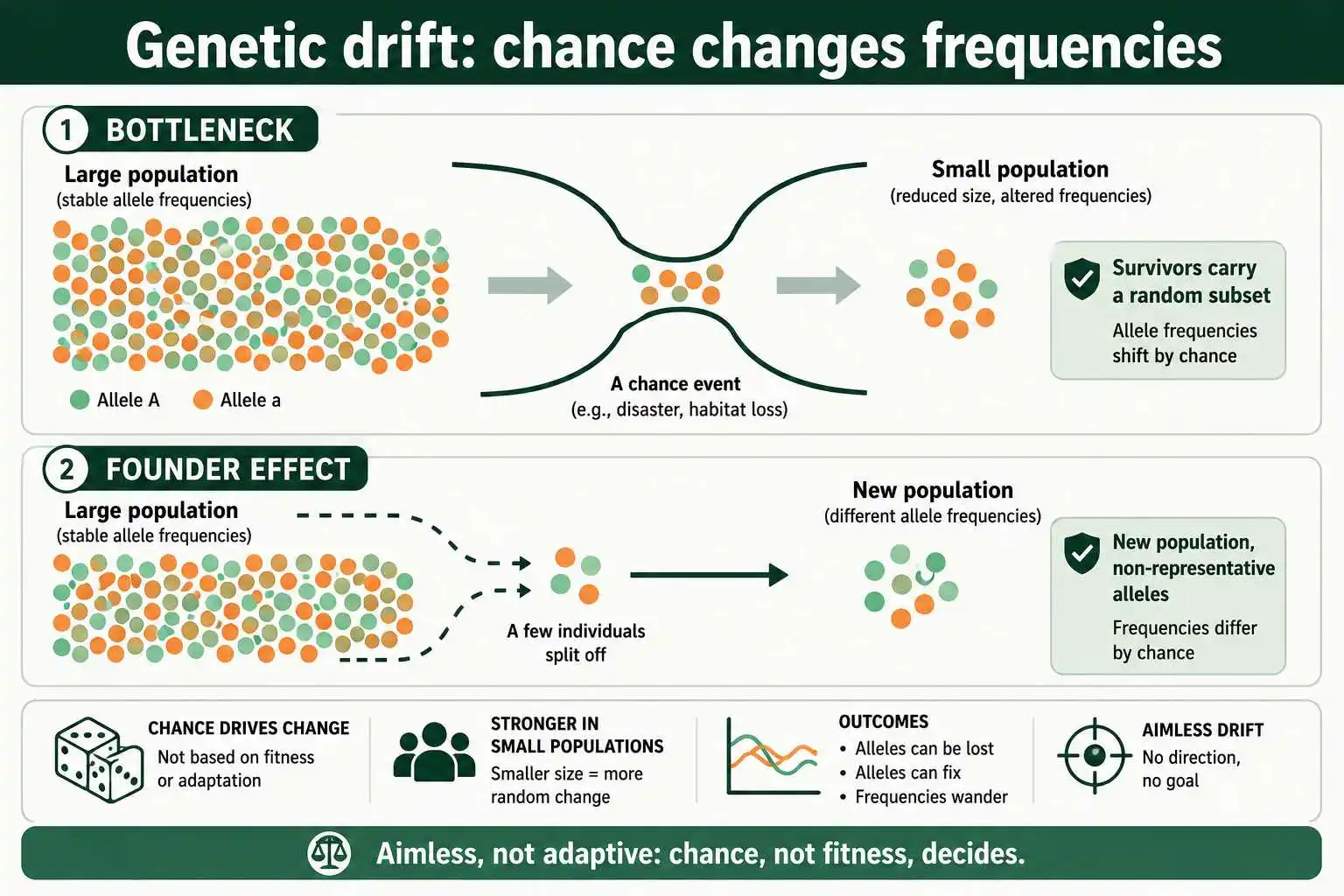
Two special cases of drift are especially important. A population bottleneck happens when a population crashes in size, so the survivors carry only a random subset of the original variation; the cheetah is a famous example, with very low genetic diversity traced to an ancient bottleneck. The founder effect occurs when a few individuals start a new population, carrying only a non-representative sample of the source population's alleles. Both dramatically alter allele frequencies through chance alone.
An important feature of drift is that it is aimless rather than adaptive. Selection improves the fit between organism and environment, but drift does no such thing; the allele that happens to increase or become fixed need not be the better one. A harmful allele can drift to high frequency in a small population, and a beneficial one can be lost, purely by chance. This is why drift is described as a non-adaptive force: it produces evolutionary change with no direction and no improvement in fitness. In conservation, this is a serious concern, because small, isolated populations can accumulate harmful alleles and lose beneficial variation through drift, reducing their long-term viability even when no selective pressure favors that outcome. The strength of drift depends on population size, being negligible in large populations and dominant in small ones, a relationship modeled directly by a genetic drift simulator.
Gene Flow
Gene flow, also called migration, is the movement of alleles between populations, and it changes allele frequencies whenever individuals move from one population to another and breed. Incoming migrants bring their alleles, and departing individuals take theirs, so both the source and receiving populations have their frequencies altered.
The effect of gene flow is generally to make populations more genetically similar to one another. When migrants from one population breed into another with different allele frequencies, they pull the receiving population's frequencies toward those of the source. Over time, ongoing gene flow tends to homogenize populations, reducing the genetic differences between them. This directly opposes the diverging effects of local selection and drift, which would otherwise make isolated populations increasingly distinct. Gene flow can also introduce entirely new alleles into a population that did not previously have them.
The amount of gene flow needed to keep populations similar is surprisingly small. Population genetics theory shows that even a few migrants exchanged between populations each generation is enough to prevent them from diverging much through drift, because a small but steady trickle of alleles continually counteracts the random local changes. This is why populations have to be quite thoroughly isolated, by distance, geography, or behavior, before they drift apart genetically. When gene flow is cut off, perhaps by a new mountain range or a human-made barrier, the previously connected populations are freed to diverge under drift and local selection, which over long periods can contribute to the formation of distinct populations and eventually new species.
A clear example is human populations connected by migration. Historically isolated groups that come into contact through migration begin to share alleles, so the genetic distinctions between them blur over generations. In conservation biology, deliberately introducing individuals from one population into another, sometimes called a genetic rescue, uses gene flow to add variation to a small, inbred population and improve its health. The degree of genetic differentiation between populations, which gene flow reduces, can be quantified with statistics like Fst, a standard way to gauge how much populations have diverged.
Mutation
Mutation is the force that creates new alleles by changing the DNA sequence, and it is the ultimate source of all genetic variation. Without mutation, there would be no alleles for the other forces to act on, so it holds a unique foundational place even though its direct effect on allele frequencies is usually slow.
Mutation disrupts equilibrium by altering allele frequencies, either introducing entirely new alleles or converting one existing allele into another. However, mutation rates are typically very low, on the order of one change per gene per many thousands or millions of copies, so the frequency shift caused by mutation in a single generation is tiny. This is why mutation, acting alone, is considered a weak force for changing allele frequencies in the short term. It rarely moves a population far from equilibrium on its own over just a few generations.
The deeper importance of mutation lies on long timescales and in its partnership with the other forces. Mutation supplies the raw genetic material, the new variants, that selection and drift then act upon. A beneficial mutation may arise rarely, but once it exists, selection can amplify it. So although mutation is a minor force for short-term frequency change, it is absolutely essential to evolution as the only process that generates new variation. Gene and genome duplication, a form of mutation, has been especially important over evolutionary history. Mutation is the slow but indispensable wellspring from which all the variation in a gene pool ultimately flows, the starting point that makes every other force possible.
Non-Random Mating: The Special Case
Non-random mating is the fifth disrupting force, and it behaves differently from the other four in a way that is essential to understand. It occurs when individuals do not pair by chance but instead choose mates based on genotype or phenotype, which alters genotype frequencies without changing allele frequencies.
This is the key distinction. The other four forces all change allele frequencies, the proportions of each allele in the gene pool. Non-random mating does not. It rearranges how the existing alleles combine into genotypes, but the overall count of each allele stays the same. The most common form is inbreeding, mating between relatives, which increases homozygosity across all genes because related individuals are more likely to share alleles. Assortative mating, where similar individuals pair, has a comparable effect for the traits involved, raising homozygotes and lowering heterozygotes relative to the Hardy-Weinberg prediction.
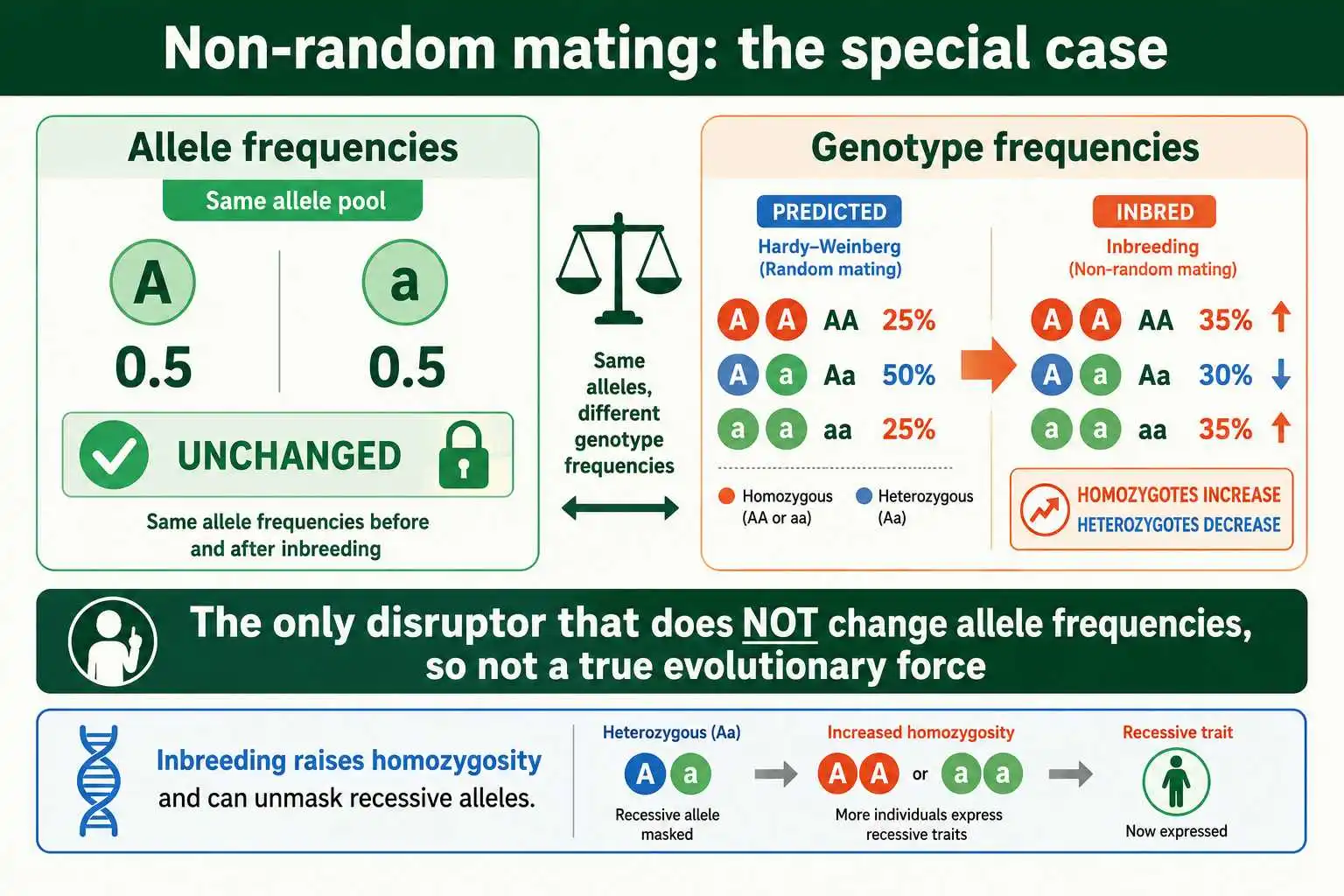
Because it leaves allele frequencies unchanged, non-random mating is technically not an evolutionary force in the strict sense, since evolution is defined as a change in allele frequencies. Yet it absolutely disrupts Hardy-Weinberg equilibrium, because the equilibrium predicts specific genotype frequencies, and non-random mating violates those. A population practicing strong inbreeding can have exactly the predicted allele frequencies while showing far more homozygotes than the p² and q² terms predict. The extent of inbreeding can be quantified with an inbreeding coefficient calculator, which measures the resulting increase in homozygosity. This makes non-random mating a disruptor of genotype equilibrium without being a driver of allele-frequency evolution.
There is a subtle way non-random mating can still influence evolution indirectly, worth noting for completeness. By changing genotype frequencies, especially by increasing homozygosity through inbreeding, non-random mating exposes more recessive alleles in the homozygous state where selection can act on them. Recessive alleles hidden in heterozygotes are shielded from selection, but inbreeding brings them together into homozygotes, making them visible to selective pressure. So while non-random mating does not itself change allele frequencies, it can alter the conditions under which selection operates, indirectly affecting how other forces play out. This interaction is one reason inbreeding can be harmful: it unmasks deleterious recessive alleles, a phenomenon known as inbreeding depression.
How the Forces Work Together
In real populations, these forces rarely act one at a time. They operate simultaneously, sometimes reinforcing and sometimes opposing one another, and the actual evolution of a population reflects their combined effect. Understanding them individually is the foundation, but appreciating their interplay is what captures real biology.
A few interactions are especially important. Selection and drift can pull in the same or opposite directions: in a small population, drift can overwhelm weak selection, so that even a slightly beneficial allele is lost by chance, while in a large population selection dominates. Gene flow opposes both local selection and drift by importing alleles and homogenizing populations, which can either rescue a population from inbreeding or swamp a locally adapted gene pool. Mutation feeds variation into the system continuously, supplying the alleles that selection and drift then sort. These combined dynamics are central to evolutionary biology and conservation, where managing small, fragmented populations requires balancing drift, gene flow, and selection at once. As one open biology text summarizes, population geneticists identified these five mechanisms precisely because each can disturb equilibrium and thereby cause evolution.
This interplay is also why detecting a deviation from Hardy-Weinberg equilibrium is the starting point rather than the end of analysis. A deviation tells you that some force is acting, but disentangling which forces, and in what combination, requires further investigation. The signature left in the genotype frequencies can hint at the cause, an excess of homozygotes suggesting inbreeding, a directional frequency change suggesting selection or gene flow, but real populations usually reflect several forces at once. Teasing them apart is the central work of population genetics, often requiring data across many genes, multiple generations, or several populations before a confident conclusion about which forces are acting can be reached. The full method for testing whether a population deviates significantly from equilibrium is covered in our guide on testing Hardy-Weinberg with chi-square.
Frequently Asked Questions
What are the five forces that disrupt Hardy-Weinberg equilibrium?
The five forces are mutation, gene flow (migration), genetic drift, non-random mating, and natural selection. Four of them, mutation, gene flow, drift, and selection, change allele frequencies, while non-random mating changes only genotype frequencies.
Which disrupting force does not change allele frequencies?
Non-random mating. It rearranges how alleles combine into genotypes, typically increasing homozygotes through inbreeding, but the overall allele frequencies stay the same. This is why it is sometimes said not to be a true evolutionary force, even though it disrupts equilibrium.
What is the difference between genetic drift and natural selection?
Natural selection changes allele frequencies in a directed way, favoring alleles that improve fitness. Genetic drift changes frequencies randomly, by chance sampling, with no regard to fitness. Selection is strongest in any population, while drift matters most in small ones.
Why is mutation considered a weak disrupting force?
Because mutation rates are very low, so mutation changes allele frequencies extremely slowly over a few generations. Its importance is as the ultimate source of new alleles, supplying the variation that selection and drift act on, rather than as a fast driver of frequency change.
Forces Behind the Frequencies
Hardy-Weinberg equilibrium is disrupted by five forces: natural selection, genetic drift, gene flow, mutation, and non-random mating. Four of them change allele frequencies and so cause evolution, with selection acting directionally toward fitness, drift acting randomly in small populations, gene flow mixing populations, and mutation slowly supplying new variation. Non-random mating stands apart, disrupting the genotype predictions while leaving allele frequencies unchanged.
These forces are not just disruptors of a tidy equation; they are the mechanisms of evolution itself, and Hardy-Weinberg equilibrium is valuable precisely because deviations from it reveal them at work. In real populations the forces act together, which is why population genetics treats a deviation as the beginning of investigation. You can generate equilibrium predictions to compare against real data with the allele frequency calculator. For an authoritative discussion of how selection, drift, and gene flow interact in natural populations, this article from Nature Education is a thorough reference.