Genetic Drift vs Natural Selection: Differences

Genetic drift and natural selection are both mechanisms of evolution that change allele frequencies, but they work in opposite ways. Natural selection is directional and non-random, favoring alleles that improve survival and reproduction. Genetic drift is random and non-directional, changing allele frequencies by pure chance, regardless of fitness. Selection builds adaptation; drift does not. The single biggest factor separating their influence is population size: selection dominates in large populations, while drift dominates in small ones.
This guide compares genetic drift and natural selection side by side, with a clear table, vivid examples, and real research data showing both forces at work. It explains how each process changes a gene pool, why they produce such different outcomes, and how they interact in real populations. A simulator can show drift operating without any selective pressure, isolating the role of pure chance. By the end, you will be able to tell these two evolutionary mechanisms apart with confidence.
The Core Difference: Direction Versus Chance
The fundamental difference is that natural selection is directional while genetic drift is random. Natural selection pushes allele frequencies in a consistent direction, toward variants that improve fitness in the current environment. Genetic drift has no direction at all, changing frequencies up or down by chance alone.
A classic analogy makes the contrast vivid. Imagine a population of frogs, some brightly colored and some drab. If herons arrive and eat the easy-to-spot bright frogs, the population becomes drabber over generations. That is natural selection: the change has a direction tied to survival, because camouflage helps frogs avoid predators. Now imagine a herd of elephants tramples through and crushes frogs at random, regardless of color. The population's color frequencies still change, but in no particular direction, because the deaths had nothing to do with fitness. That is genetic drift.

Both processes are forms of evolution, since both change allele frequencies across generations. The crucial distinction is the cause of the change. Under natural selection, allele frequencies shift because certain alleles confer a survival or reproductive advantage. Under genetic drift, they shift purely because of random sampling, the luck of which individuals happen to survive and reproduce. This is why selection is often called a non-random process and drift a random one. Understanding drift on its own is the foundation for this comparison, covered fully in our guide on what genetic drift is.
Genetic Drift vs Natural Selection: Comparison Table
The clearest way to grasp the differences is to see the two mechanisms side by side. The table below summarizes the key contrasts across the features that matter most.
| Feature | Natural selection | Genetic drift |
|---|---|---|
| Direction | Directional, predictable | Random, unpredictable |
| Driven by | Fitness and environment | Chance sampling |
| Relationship to fitness | Favors beneficial alleles | Ignores fitness entirely |
| Effect on adaptation | Increases adaptation | Does not increase adaptation |
| Strongest in | Large populations | Small populations |
| Effect on diversity | Can increase or decrease it | Tends to reduce it |
| Outcome | Better-adapted populations | Random allele fixation or loss |
| Example | Peppered moth color shift | Founder effect, bottleneck |
Each row highlights a genuine, testable difference. Natural selection is the only one of the two that consistently produces adaptation, the fit between organism and environment. Genetic drift, by contrast, changes a population's genetics without improving its fit to anything. The population-size row is especially important, because it explains when each force dominates. The rest of this guide unpacks these contrasts and shows how the two forces play out together in real populations.
How Each Process Changes a Gene Pool
Natural selection changes a gene pool by sorting alleles according to their effect on survival and reproduction. Alleles that help an organism survive and reproduce become more common, because their carriers leave more offspring. Alleles that hurt fitness become rarer, because their carriers leave fewer offspring. This sorting is consistent and repeatable.
The mechanism of selection is differential reproductive success. In any environment, some genotypes produce more surviving offspring than others, and those genotypes pass on more copies of their alleles. Over generations, the beneficial alleles accumulate and the population becomes better adapted. Because the same environmental pressure produces the same directional change, natural selection is predictable: if dark coloration helps moths hide from predators, the dark allele will tend to increase wherever those predators hunt by sight.
Genetic drift changes a gene pool through random sampling error in reproduction. Each generation is a finite sample of the previous generation's alleles, and small samples rarely match the original frequencies exactly. The mechanism here is luck, not fitness. An allele might increase simply because its carriers happened to reproduce more, with no advantage involved, or it might be lost because its few carriers happened not to reproduce. Over time this random walk can carry an allele to fixation at 100 percent or to complete loss, with no relationship to whether the allele was useful. The two mechanisms thus reshape the gene pool through entirely different logics: selection by merit, drift by chance.
A simple numerical contrast captures the difference. Suppose an allele starts at a frequency of 50 percent in a population. Under strong natural selection favoring that allele, its frequency might rise steadily and predictably, perhaps to 55 percent, then 60, then 65 percent over successive generations, climbing in a consistent direction because each generation the fitter carriers leave more offspring. Under pure drift, the same allele might wander to 47 percent, then 52, then 44 percent, bouncing around with no trend, because each generation is just a random sample. Run the selection scenario again and the allele rises again, but run the drift scenario again and it might wander the opposite way. That repeatability under selection versus unpredictability under drift is the practical fingerprint of the two forces.
Why Population Size Is the Deciding Factor
Population size determines which force dominates, because the strength of genetic drift is inversely related to population size. Drift is powerful in small populations and weak in large ones, while natural selection works effectively across population sizes and is relatively stronger when populations are large.
The reason traces to sampling, the same coin-flip logic that governs all of drift. In a large population, each generation is a huge sample of the gene pool, so random fluctuations average out and frequencies stay stable, leaving selection free to act on even small fitness differences. In a small population, each generation is a tiny sample, so chance can swing frequencies dramatically, often overwhelming the gentle push of selection. A slightly beneficial allele that selection would favor can be lost by sheer bad luck in a small population, before selection has a chance to act.
This is why the relevant number is the effective population size, not the raw headcount. The effective population size is the number of individuals actually contributing genes to the next generation, which is usually smaller than the total because not everyone breeds. Drift acts according to this effective size. The practical consequence is a clear rule of thumb: in large populations, selection tends to win and adaptation proceeds efficiently; in small populations, drift can dominate, and chance can override fitness. This interplay between size and the two forces is central to evolutionary biology, and the effective size concept is covered in our guide on effective population size.
Real Data: Both Forces at Work
Real studies show that drift and selection often operate together, and sometimes drift is the bigger player even when selection is present. A striking example comes from a 60-year study of the scarlet tiger moth, Callimorpha dominula, which tracked the frequency of a color-pattern genotype over time.
The analysis of this long time series found that most of the variation in the genotype's frequency was the result of genetic drift, not selection. Selection was acting, but its measurable effect was small, and mean fitness barely increased over the decades. This was a population large enough to study in detail, yet chance still explained the bulk of the genetic change. The study is a reminder that drift is not just a small-population curiosity confined to founder events and bottlenecks; it operates continuously, and its contribution can rival or exceed that of selection even in ongoing natural populations.
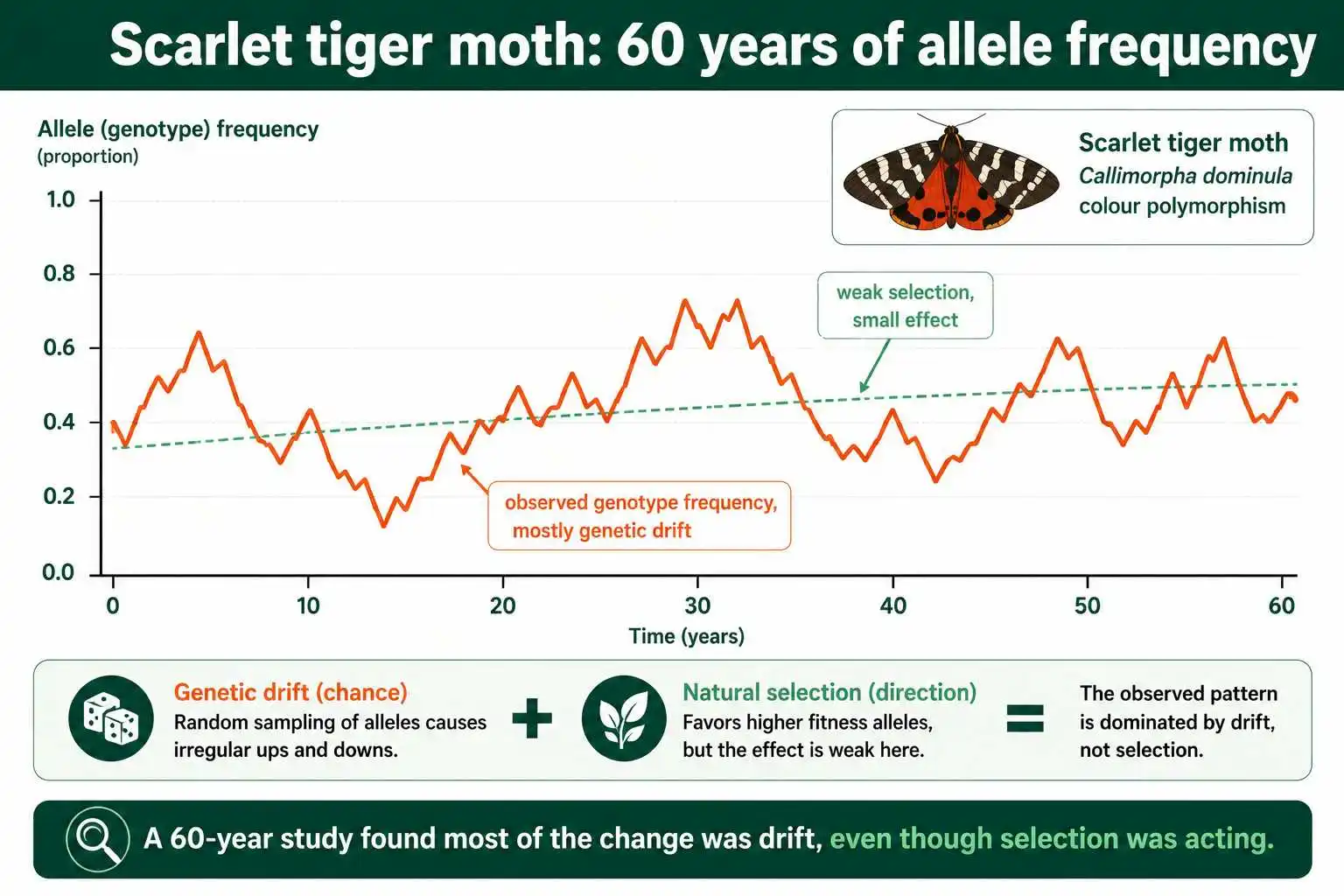
This difficulty of separating the two forces is a genuine challenge in evolutionary biology. When a population's allele frequencies change, it is often hard to tell how much was drift and how much was selection. Researchers have developed statistical methods to distinguish them, such as comparing DNA sequences among related strains or analyzing quantitative trait loci, but the question is rarely simple. The honest scientific picture is that most real evolution reflects a mix of both forces, with their relative importance depending heavily on population size, the strength of selection, and the specific gene in question. Pure selection and pure drift are the endpoints of a spectrum that real populations fall somewhere along.
Do Both Cause Evolution and Speciation?
Both genetic drift and natural selection are genuine mechanisms of evolution, because both change allele frequencies in a population over time. Evolution is defined as a change in allele frequencies across generations, and drift and selection both produce that change, just by different means.
Both can also contribute to speciation, the formation of new species. Natural selection drives speciation when populations in different environments adapt to their local conditions, accumulating genetic differences until they can no longer interbreed. Genetic drift drives speciation when isolated populations, especially small ones, accumulate random genetic differences over time. A population split by a geographic barrier can drift in different directions on each side, and those random divergences can eventually contribute to reproductive isolation. Often the two work together, with drift and selection jointly pushing isolated populations apart.
The key difference in their evolutionary roles is adaptation. Natural selection is the only mechanism that consistently produces adaptation, the apparent design that fits organisms to their environments. It multiplies beneficial mutations and eliminates harmful ones, accounting for the organized complexity of living things. Genetic drift produces evolutionary change without adaptation; it alters a population's genetics in directions unrelated to fitness. So while both are evolution, only selection explains why organisms appear so well suited to their ways of life. Drift explains much of the random genetic variation among populations that selection cannot account for, including many of the differences revealed by molecular data.
This adaptation contrast is why the two forces leave different fingerprints in the genome. Natural selection leaves signatures of adaptation, such as a beneficial allele sweeping to high frequency along with the DNA around it, a pattern called a selective sweep. Genetic drift leaves a more uniform, directionless pattern of variation across the genome, with no particular allele favored. Population geneticists exploit exactly these differences to scan genomes for regions shaped by selection against the background hum of drift, which is how researchers identify the specific genes underlying adaptations like disease resistance or altitude tolerance.
How the Two Forces Interact
In real populations, genetic drift and natural selection act simultaneously, sometimes reinforcing each other and sometimes opposing each other. They are not alternatives that take turns; they operate at the same time on the same gene pool, and the net result reflects their combined influence.
Their interaction often comes down to a tug of war set by population size and selection strength. When selection is strong and the population is large, selection usually prevails, efficiently increasing beneficial alleles. When selection is weak or the population is small, drift can override it, so that even a beneficial allele may be lost or a harmful one fixed by chance. A nearly neutral allele, one with only a tiny fitness effect, is especially subject to drift, because the random fluctuations easily swamp the faint selective signal. This is the insight behind the nearly neutral theory of molecular evolution, which holds that much genetic change is governed by the balance between weak selection and drift.
Gene flow and mutation add further complexity, but the drift-selection balance is the central dynamic. Understanding it explains many real patterns: why small, isolated populations can fix harmful alleles, why large populations adapt more efficiently, and why molecular evolution often looks more random than the adaptive story alone would predict. The two forces are best understood not as rivals but as partners that jointly shape every population's genetics, with their relative weight shifting according to circumstances. This combined action, alongside mutation and gene flow, is the full picture of how allele frequencies change, explored in our guide on what disrupts Hardy-Weinberg equilibrium.
How to Tell Which Force Is Responsible
Telling whether genetic drift or natural selection caused a change is one of the harder tasks in evolutionary biology, but several clues help distinguish them. The distinction matters because attributing a change to the wrong force leads to wrong conclusions about why a population evolved.
The strongest clue is whether the change is correlated with fitness or environment. If an allele increases specifically where it provides an advantage, and the increase tracks an environmental pressure, that pattern points to natural selection. The peppered moth is a clear case: the dark allele rose precisely where industrial soot darkened tree bark and made dark moths better camouflaged, a tight link between the allele, the environment, and survival. When a change shows no such link to fitness or environment, drift becomes the more likely explanation.
Population size and repeatability offer further clues. Drift is more plausible in small populations, where chance is powerful, and its hallmark is unpredictability, since independent populations drift in different random directions. Selection, by contrast, is repeatable: the same environmental pressure produces the same directional change in separate populations, so parallel evolution across populations strongly suggests selection rather than drift. Scientists also use formal statistical tests, comparing DNA sequences among related lineages or analyzing patterns across many genes, to detect the signature of selection against a background of drift. Even with these tools, the answer is often a matter of degree rather than a clean verdict, because both forces usually contribute.
Common Misconceptions About the Two Forces
Several misconceptions cloud the comparison between drift and selection, and clearing them up sharpens understanding. These errors are common among students and worth addressing directly.
The first misconception is that genetic drift only matters in tiny or endangered populations. In reality, drift operates in every finite population, all the time, and as the scarlet tiger moth study showed, it can account for most genetic change even in a sizable population. Drift is weaker in large populations, but never absent. The second misconception is that natural selection is random. Selection is the opposite of random; it is the consistent, non-random sorting of alleles by fitness. The randomness in evolution comes from mutation, which generates variants blindly, and from drift, which shuffles them by chance, not from selection.
A third misconception is that drift and selection are mutually exclusive, as if a change must be caused by one or the other. In fact they act together constantly, and most real evolutionary change reflects a blend of both. A fourth is the belief that drift always reduces fitness while selection always increases it. Selection does tend to raise mean fitness, but drift is indifferent to fitness rather than always harmful; by chance it can occasionally fix a beneficial allele or remove a harmful one, though on average it erodes the diversity a population needs. Recognizing that drift is genuinely directionless, neither helpful nor harmful by design, is the key to understanding how it differs from the purposeful sorting of selection.
Frequently Asked Questions
What is the main difference between genetic drift and natural selection?
The main difference is direction. Natural selection changes allele frequencies in a predictable direction, favoring alleles that improve fitness. Genetic drift changes allele frequencies randomly, by chance, with no regard for fitness. Selection builds adaptation, while drift does not.
Which is stronger, genetic drift or natural selection?
It depends on population size. Genetic drift is stronger in small populations, where chance can override fitness. Natural selection is relatively stronger in large populations, where random fluctuations average out and even small fitness advantages can drive change.
Do genetic drift and natural selection both cause evolution?
Yes, both are mechanisms of evolution because both change allele frequencies over generations. The difference is that natural selection produces adaptation by favoring beneficial alleles, while genetic drift changes a population's genetics randomly, without improving its fit to the environment.
Can genetic drift and natural selection happen at the same time?
Yes, they act simultaneously on the same population. Their combined effect depends on population size and the strength of selection. In large populations selection tends to dominate, while in small populations drift can override selection, even eliminating a beneficial allele by chance.
Telling the Two Forces Apart
Genetic drift and natural selection are both engines of evolution, but they could hardly be more different in how they work. Natural selection is directional and non-random, sorting alleles by fitness to build adaptation. Genetic drift is random and non-directional, shuffling allele frequencies by chance with no regard for fitness, and tending to erode genetic diversity. The deciding factor between them is population size: selection dominates when populations are large, and drift dominates when they are small.
The deeper truth is that real populations experience both at once, in a balance set by their size and the strength of selection. As the scarlet tiger moth study showed, chance can account for the bulk of genetic change even when selection is present. Evolution, in practice, is the combined product of merit and luck. You can isolate the role of pure chance, with no selection at all, using the genetic drift simulator, a useful way to see what drift does on its own. For an authoritative account of how drift relates to population size, this overview from Britannica is a clear reference, and the scarlet tiger moth study documents both forces acting over six decades.