Mutation Rate vs Substitution Rate

Mutation rate and substitution rate are different things that people constantly mix up. The mutation rate is how fast new mutations arise in an individual. The substitution rate is how fast those mutations spread and become fixed across an entire population or species. They are linked by one of the most elegant results in genetics: for neutral mutations, the two rates are exactly equal.
This guide separates the two clearly and then shows how they connect. It covers what each measures, the neutral-theory result that the substitution rate equals the mutation rate, why selection breaks that equality, and why the whole idea underpins the molecular clock. For the first quantity, our explainer on what mutation rate is covers it in full.
Two Different Rates
The cleanest way to start is with what each rate is about. Mutation rate is an individual-level quantity; substitution rate is a population-level one.
The mutation rate is the frequency at which new mutations appear, usually per base pair per generation. It describes events in single individuals: a new change arising in an egg, a sperm, or a dividing cell. The human germline mutation rate of about 1.2 x 10^-8 per base per generation is a mutation rate. It counts mutations as they originate, before anything else happens to them.
The substitution rate is the frequency at which mutations become fixed in a population, meaning they spread until essentially every individual carries them. A substitution is the end of a long journey: a mutation arises in one individual, then over many generations rises in frequency until it replaces the previous version across the whole population. The substitution rate counts only the mutations that complete that journey, per unit time.
So the two rates sit at opposite ends of a mutation's life. The mutation rate counts mutations at birth; the substitution rate counts the few that eventually take over. Most mutations never make it, which is why you might expect the substitution rate to be far lower than the mutation rate. For neutral mutations, remarkably, it is not.
The Neutral Result: Substitution Rate Equals Mutation Rate
The central result, from Motoo Kimura's neutral theory, is that for selectively neutral mutations the substitution rate equals the mutation rate. Written as an equation, k = u, where k is the substitution rate and u is the mutation rate. This looks surprising until you see the derivation, which is short and beautiful.
Consider a population of N individuals, and focus on neutral mutations at some site. Each generation, the number of new neutral mutations entering the population is the mutation rate times the number of gene copies, which is 2Nu in a diploid population. That is the supply of new mutations.
Now the fate of each. Because the mutations are neutral, none has any selective advantage, so each new mutation's chance of eventually fixing is simply its frequency in the population, which for a brand-new mutation is 1 divided by 2N. That is the fixation probability.
Multiply the two. The substitution rate is the number of new mutations per generation times the probability each one fixes: 2Nu times 1/(2N). The population size cancels completely, leaving k = u. The substitution rate equals the mutation rate, regardless of population size. Kimura published this result in his 1968 paper in Nature, and it became the mathematical backbone of the neutral theory of molecular evolution.
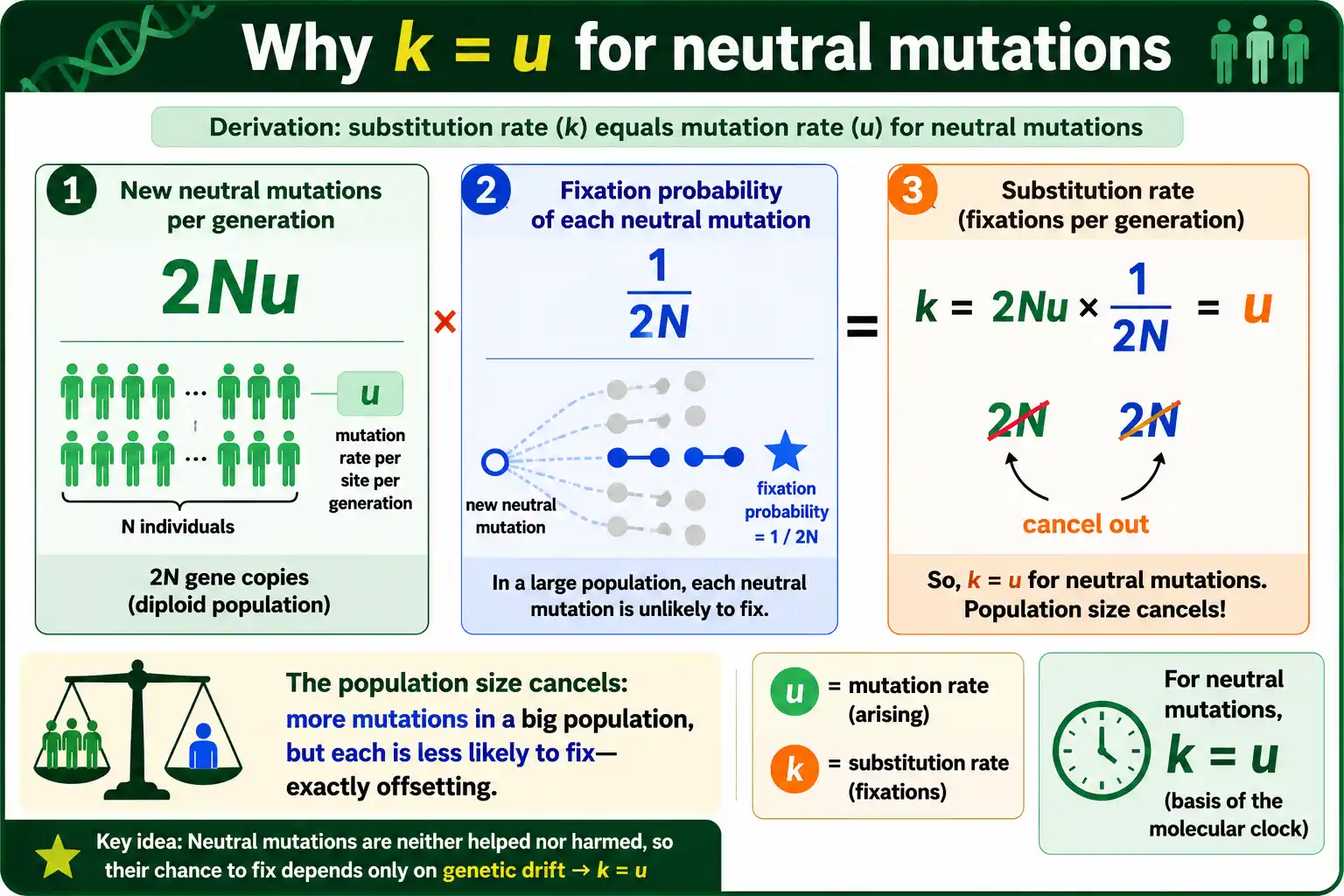
Why the Population Size Cancels
The cancellation is the heart of the result, and it is worth dwelling on because it is so counterintuitive. Larger populations generate more mutations but fix each one less easily, and the two effects exactly offset.
In a big population, there are more individuals, so more new neutral mutations arise each generation, the supply term 2Nu grows. But in that same big population, any single new mutation starts at a smaller frequency, 1/(2N), so it is less likely to drift to fixation. More mutations enter, but each is less likely to win. The two effects are precise mirror images, so their product does not depend on N at all.
This is why the neutral substitution rate is set entirely by the mutation rate, not by population size. It is also why genetic drift is the engine here: in the absence of selection, the mutations that fix do so purely by the random sampling of drift, and drift fixes neutral alleles at a rate that depends only on how fast they arise. The connection to drift and population size is developed in our guides on genetic drift and effective population size, which explain the fixation side of this story in detail.
When Selection Breaks the Equality
The equality k = u holds only for neutral mutations. When selection acts, the substitution rate departs from the mutation rate, and the direction of that departure is itself informative.
For advantageous mutations, selection raises the fixation probability above the neutral 1/(2N), so beneficial mutations fix more often than chance, and the substitution rate exceeds the mutation rate: k is greater than u. Positively selected sequences evolve faster than neutral ones.
For deleterious mutations, selection lowers the fixation probability below 1/(2N), so harmful mutations are usually removed before they can spread, and the substitution rate falls below the mutation rate: k is less than u. Sequences under purifying, or negative, selection evolve more slowly than neutral ones, because most changes are weeded out.
This gives a powerful test. By comparing the actual substitution rate at a sequence to the neutral expectation, biologists can detect selection. A region evolving faster than the neutral mutation rate is a candidate for positive selection; one evolving slower is under purifying selection. The neutral case, where k equals u, is the null hypothesis against which selection is measured, which is why the neutral theory is often called the null model of molecular evolution.
A Worked Example of the Cancellation
Concrete numbers make the cancellation click. Compare two neutral populations, one small and one large, at a site with a mutation rate u of 1 x 10^-8 per generation.
Take the small population first, with 1,000 diploid individuals. New neutral mutations per generation are 2Nu, which is 2 times 1,000 times 1 x 10^-8, or 2 x 10^-5 mutations. Each has a fixation probability of 1/(2N), which is 1/2,000, or 5 x 10^-4. The substitution rate is their product: 2 x 10^-5 times 5 x 10^-4, which is 1 x 10^-8. That equals u.
Now the large population, with 1,000,000 diploid individuals. New mutations per generation are 2 times 1,000,000 times 1 x 10^-8, or 2 x 10^-2, a thousand times more than the small population. But each mutation's fixation probability is now 1/2,000,000, or 5 x 10^-7, a thousand times smaller. The product is 2 x 10^-2 times 5 x 10^-7, which is again 1 x 10^-8. Identical to the small population, and equal to u.
The thousandfold larger supply of mutations in the big population is exactly canceled by their thousandfold lower chance of fixing. Whatever the population size, the neutral substitution rate lands on the mutation rate. This is the cancellation working in plain arithmetic, and it is why the molecular clock does not depend on how big a population was.
Detecting Selection With dN/dS
The most widely used application of the substitution-equals-mutation idea is a test that compares two kinds of substitutions within protein-coding genes. It turns the neutral baseline into a practical selection detector.
Mutations in protein-coding DNA come in two flavors. Synonymous changes do not alter the encoded amino acid, so they are usually close to neutral. Nonsynonymous changes do alter the amino acid, so selection can act on them. Counting the substitution rate of each, the nonsynonymous rate dN and the synonymous rate dS, and taking their ratio gives the dN/dS statistic.
The interpretation flows straight from the neutral result. If a gene evolves neutrally, nonsynonymous and synonymous changes fix at the same rate, so dN/dS is about one. If purifying selection is removing amino-acid changes, dN is suppressed and dN/dS falls below one, the usual case for functional genes. If positive selection is driving amino-acid changes to fixation, dN exceeds dS and dN/dS rises above one, a signal of adaptive evolution. Because synonymous sites act as a built-in neutral yardstick, the test reads selection directly off the comparison, and it is one of the most common ways researchers scan genomes for genes under selection. It is the neutral theory turned into a routine tool.
The Molecular Clock
The equality of substitution and mutation rates for neutral sites is what makes the molecular clock work. The clock is the observation that molecular differences between species accumulate at a roughly steady rate over time, letting genetic distance estimate divergence time.
The logic follows directly. If neutral mutations fix at a constant rate equal to the mutation rate, then the number of substitutions separating two species grows steadily with the time since they diverged. Count the differences, divide by the rate, and you get the time, the basis of dating evolutionary splits from DNA. The clocklike regularity was first noticed by Emile Zuckerkandl and Linus Pauling in the early 1960s, and Kimura's neutral theory later explained why it should exist.
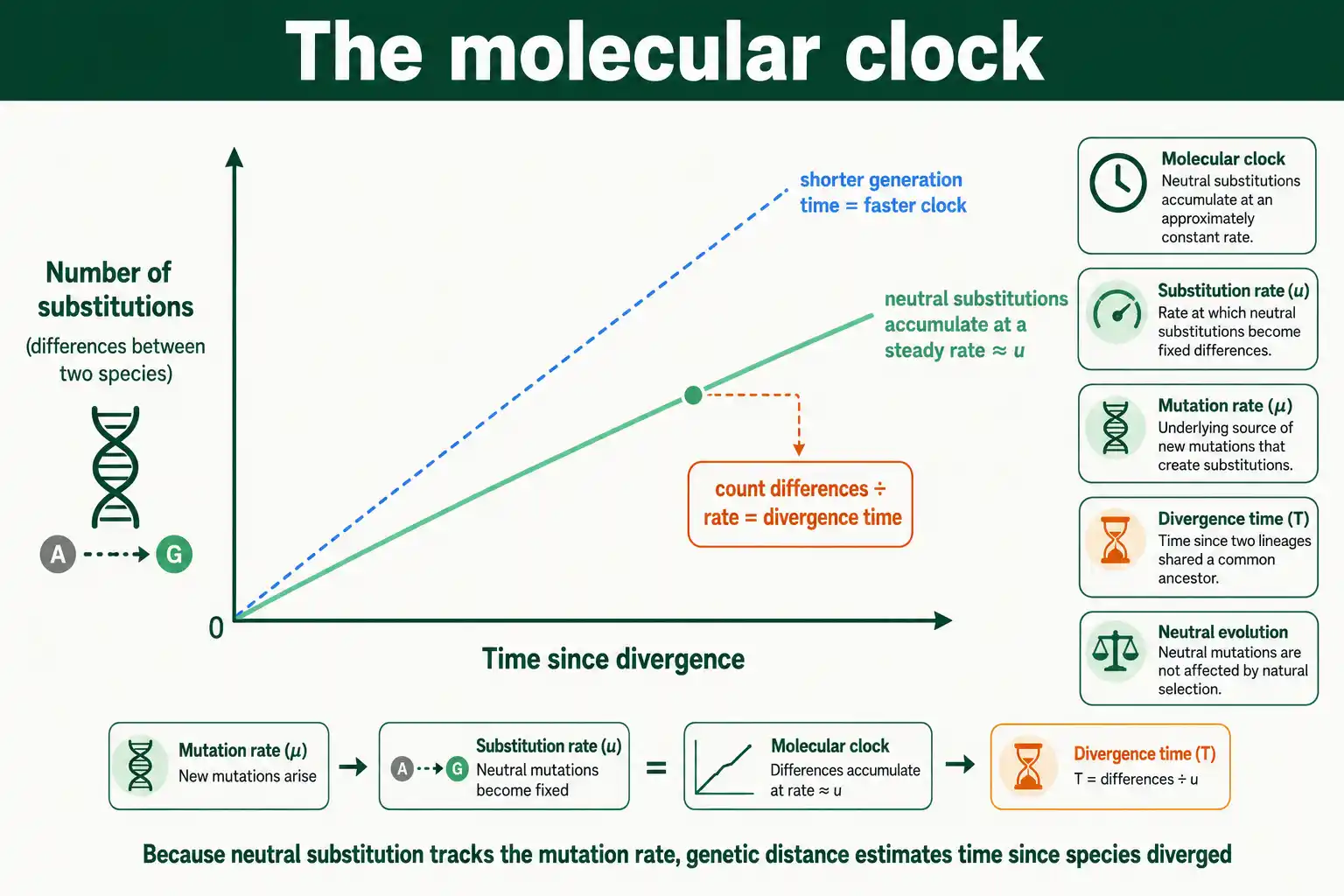
The clock is not perfect. Generation times differ between lineages, mutation rates themselves vary, and selection on some sequences disturbs the regularity, so molecular clocks must be calibrated and used carefully. But the core insight stands: because neutral substitution tracks the mutation rate, DNA carries a rough record of elapsed time, which is the foundation of molecular dating across the tree of life.
One well-documented wrinkle is the generation-time effect. Species with shorter generations copy their DNA more often per unit of absolute time, so they tend to accumulate substitutions faster per year even when the per-generation mutation rate is similar. This is part of why the molecular clock runs at different speeds in different lineages, with small, fast-breeding rodents ticking faster than large, slow-breeding primates. Modern molecular dating handles this with relaxed clocks that allow the rate to vary across the tree, calibrated against fossil dates, rather than assuming a single universal tick. The neutral result still anchors all of it, because without the equality of substitution and mutation rates there would be no clock to relax in the first place.
Side by Side
The table sums up the contrast.
| Feature | Mutation rate (u) | Substitution rate (k) |
|---|---|---|
| Level | Individual | Population or species |
| Counts | New mutations as they arise | Mutations that reach fixation |
| Typical symbol | u or µ | k |
| For neutral mutations | Equal to k | Equal to u |
| Under positive selection | Unchanged | Higher than u (k greater than u) |
| Under purifying selection | Unchanged | Lower than u (k less than u) |
| Main use | Disease risk, raw variation | Molecular clock, detecting selection |
The key row is the neutral one. The mutation rate is a property of molecular biology, set by replication fidelity and repair. The substitution rate is a property of evolution, set by mutation plus the fate of those mutations under drift and selection. They coincide only when selection is absent, and that coincidence is the single fact that makes the molecular clock possible.
Frequently Asked Questions
Is the substitution rate the same as the mutation rate?
Only for selectively neutral mutations. The neutral theory shows that for neutral sites the substitution rate equals the mutation rate, k = u, because the population size cancels out of the math. For mutations under selection the two differ: positive selection makes the substitution rate exceed the mutation rate, while purifying selection makes it fall below.
Why does the substitution rate equal the mutation rate for neutral alleles?
Because two effects of population size cancel. Each generation a population of N diploid individuals generates 2Nu new neutral mutations, and each has a fixation probability of 1/(2N). Multiplying gives a substitution rate of u, with N canceling entirely. So neutral mutations fix at a rate set only by how fast they arise, independent of population size.
The Same Coin, Two Faces
Mutation rate and substitution rate describe a mutation at two stages of its life: arising in an individual, and fixing across a population. For neutral mutations the two rates are equal, k = u, because the larger supply of mutations in a big population is exactly offset by each one's lower chance of fixing. That cancellation, from Kimura's neutral theory, is one of the most elegant results in genetics.
Selection breaks the equality in a readable way: faster-than-neutral evolution signals positive selection, slower-than-neutral signals purifying selection, and the neutral case is the null model both are measured against. The same equality is what lets neutral substitutions act as a molecular clock for dating evolutionary history. To explore the fixation side of this story, where drift turns arising mutations into substitutions, you can model how a given mutation rate plays out across a population, and our guide on what affects the mutation rate covers what sets u in the first place.