Why Recessive Traits Don't Disappear (Explained)

Recessive traits do not disappear because dominance describes how an allele is expressed, not how common it is. A dominant allele has no built-in tendency to become more frequent, and a recessive allele has no tendency to vanish. Most copies of a recessive allele are hidden inside healthy heterozygous carriers, where natural selection cannot act on them, so the allele persists from generation to generation. This is exactly what the Hardy-Weinberg principle proved in 1908: in the absence of evolutionary forces, allele frequencies stay constant, no matter which allele is dominant.
It seems intuitive that a dominant allele should gradually take over and a recessive one fade away, yet this does not happen, and the reason reveals something fundamental about genetics. This guide explains why recessive traits persist, debunking the common misconception step by step: why dominance is not the same as frequency, how carriers shelter recessive alleles, and what the Hardy-Weinberg principle says about allele frequencies over time. The ideas are conceptual and worth understanding directly, though the underlying frequencies can be explored with a calculator.
The Misconception: Dominant Means Winning
The belief that recessive traits should disappear comes from a natural but mistaken intuition about the word dominant. Because a dominant allele masks a recessive one in a heterozygote, it is easy to assume the dominant allele is somehow stronger or winning, and that over time it should crowd the recessive allele out of the population.
This intuition is so common that it troubled early geneticists too. After Mendel's work was rediscovered around 1900, some scientists genuinely argued that dominant traits should increase in frequency and recessive traits should decrease, eventually disappearing. They reasoned that since three-quarters of the offspring of two heterozygotes show the dominant trait, the dominant version should steadily come to dominate the whole population. It was a serious scientific question, not just a student's confusion.
The mistake lies in conflating two completely different things: how an allele is expressed and how common it is. Dominance is about expression, which allele's effect shows up in a heterozygote. Frequency is about how many copies of each allele exist in the population. These are independent. A dominant allele can be rare, and a recessive allele can be common. The ratio of dominant to recessive offspring in a single cross says nothing about whether the dominant allele is becoming more frequent in the population over generations. Untangling this confusion is the key to understanding why recessive traits persist, and it was the precise issue that Hardy and Weinberg resolved.
Dominance Is Not Frequency
The single most important idea for dispelling the myth is that dominance and frequency are unrelated. An allele's dominance tells you only what happens when it is paired with a different allele in one individual; it tells you nothing about how often the allele appears across the whole population.
Consider a clear real-world example. Polydactyly, the condition of having extra fingers or toes, is caused by a dominant allele, yet it is quite rare. Most people have the recessive genotype for the normal number of digits, so here the recessive condition is overwhelmingly common and the dominant one is rare. This single example shatters the intuition that dominant equals common. If dominance forced an allele to increase, polydactyly would be widespread, but it is not, because dominance has no such power.

The reverse is just as true: many recessive alleles are extremely common in populations. Whether an allele is frequent or rare depends on the population's history, including past selection, genetic drift, mutation, and migration, never on its dominance relationship. A dominant allele does not get a reproductive head start simply for being dominant. Once you separate these two ideas firmly, dominance for expression, frequency for prevalence, the myth loses its foundation. The distinction between an allele's effect and its abundance is rooted in the difference between genotype and phenotype, which underlies the whole confusion.
What Hardy and Weinberg Proved
In 1908, Godfrey Hardy and Wilhelm Weinberg independently settled the question with a simple but powerful piece of reasoning. They showed mathematically that, in the absence of any disturbing force, allele frequencies do not change from one generation to the next, regardless of which allele is dominant.
Their insight was that the reshuffling of alleles during reproduction does not, by itself, alter their proportions. If a population starts with a certain frequency of an allele, and nothing disturbs it, the next generation will have exactly the same frequency. Dominance affects which trait you see in a heterozygote, but it has no influence on the underlying allele frequencies. The dominant allele does not convert recessive alleles into copies of itself; it simply masks them in heterozygotes while leaving their numbers untouched. This directly refuted the argument that dominant traits should increase.
This principle, now called Hardy-Weinberg equilibrium, became the foundation of population genetics. It establishes a crucial baseline: a population that is not being acted on by evolutionary forces does not change its allele frequencies at all. Recessive alleles stay at their existing frequency indefinitely under these conditions, neither increasing nor decreasing. The persistence of recessive traits is therefore not a puzzle requiring special explanation; it is the default expectation once you understand that inheritance alone does not change allele frequencies. The full statement of this principle is covered in our guide on what Hardy-Weinberg equilibrium is.
It is worth appreciating how counterintuitive this result was at the time, and how elegantly simple the resolution turned out to be. The scientists who expected dominant traits to take over were reasoning by analogy with a contest, as if alleles competed and the stronger won. Hardy and Weinberg showed there is no contest at all in the absence of selection. Each allele is simply copied and passed on in proportion to how common it already is, like drawing colored balls from a bag and replacing them in the same proportions. The proportions do not drift just because one color is labeled dominant. This reframing, from competition to neutral sampling, is what dissolved decades of confusion, and it is why the principle is considered one of the founding insights of population genetics.
How Carriers Shelter Recessive Alleles
Even when there is selection against a recessive trait, the allele still resists disappearing, and the reason is one of the most important ideas in population genetics: recessive alleles hide in heterozygous carriers. A carrier has one recessive allele and one dominant allele, shows the dominant phenotype, and is completely healthy, yet still carries and transmits the recessive allele.
This sheltering effect is dramatic for rare alleles. When a recessive allele is uncommon, the vast majority of its copies exist in heterozygous carriers rather than in affected homozygous individuals. The mathematics makes this precise: the frequency of affected homozygotes is q², while the frequency of carriers is 2pq, and when q is small, q² is far smaller than 2pq. For an allele with a frequency of 0.01, only 0.0001 of the population is affected, but about 0.02 are carriers, two hundred times as many. Almost every copy of the allele is tucked away in a healthy carrier.

This is why selection struggles to eliminate recessive alleles. Natural selection can only act on phenotypes it can see, and the recessive allele is invisible in carriers. Selection can remove affected homozygotes, but it cannot touch the much larger reservoir of alleles hidden in carriers. As the allele becomes rarer, an ever-greater proportion of its copies sit in carriers, so selection becomes less and less able to reduce it further. The allele can persist almost indefinitely at low frequency, sheltered in carriers who pass it quietly to the next generation. This carrier reservoir is quantified in our guide on Hardy-Weinberg carrier frequency.
Why Selection Slows Down as Alleles Get Rare
A deeper consequence of the carrier effect is that selection against a recessive allele becomes progressively weaker as the allele gets rarer. This explains why even harmful recessive alleles, which actively reduce the fitness of affected individuals, are not driven to extinction.
The logic follows from the carrier sheltering. When a recessive allele is common, a meaningful fraction of the population is homozygous and affected, so selection removes a substantial number of alleles each generation. But as selection lowers the allele frequency, the proportion of copies exposed in affected homozygotes shrinks rapidly, because q² falls much faster than q. At low frequencies, almost all the alleles are in carriers, untouched by selection, and only a tiny fraction are exposed in affected individuals each generation. The rate of removal slows to a crawl.
The result is that selection against a recessive allele shows strongly diminishing returns. It can reduce a common harmful allele relatively quickly, but pushing it from rare toward truly absent takes enormous numbers of generations, because each generation exposes so few copies. In practice, this means harmful recessive alleles persist in populations at low frequencies essentially indefinitely. Add in a steady trickle of new copies from mutation, and an equilibrium is reached where the rare alleles removed by selection are replaced by new mutations, holding the frequency stable. The allele never disappears; it settles at a low, persistent level.
This mutation-selection balance is a real and measurable phenomenon for many genetic conditions. The frequency at which a harmful recessive allele settles depends on how strongly selection acts against the affected homozygotes and on the rate at which new copies arise by mutation. A more harmful allele settles at a lower frequency, and a higher mutation rate pushes the balance point higher, but in every case the allele reaches a stable, nonzero level rather than vanishing. This explains why recessive genetic disorders have characteristic, fairly stable frequencies in populations over long periods, rather than steadily declining toward zero as the naive disappearing-trait intuition would predict.
When Recessive Alleles Do Change Frequency
To be precise, recessive alleles can change frequency, and even decline, but only when an evolutionary force acts, never from dominance alone. Understanding what can change a recessive allele's frequency completes the picture and clarifies exactly what the myth gets wrong.
The forces that change allele frequencies are the familiar ones that violate Hardy-Weinberg conditions: natural selection, genetic drift, gene flow, mutation, and non-random mating. Selection against affected individuals can slowly reduce a recessive allele, though, as we have seen, with sharply diminishing returns at low frequency. Genetic drift can change a recessive allele's frequency randomly in a small population, sometimes even eliminating it by chance, or conversely increasing it. Gene flow can add or remove copies through migration. So recessive alleles are not frozen forever; they respond to real evolutionary pressures.
The crucial point is that none of these changes happen because the allele is recessive. Dominance is never the cause. A recessive allele declines only if a force like selection or drift acts to reduce it, exactly as a dominant allele would decline under the same force. In fact, dominance relationships can even affect how quickly selection works, since a rare advantageous dominant allele spreads faster than a rare advantageous recessive one, precisely because the dominant allele is visible to selection in heterozygotes while the recessive one is hidden. But this is selection doing the work, not dominance. The forces that actually change frequencies are detailed in our guide on what disrupts Hardy-Weinberg equilibrium.
A Related Myth: Skipping Generations
A close cousin of the disappearing-trait myth is the idea that recessive traits "skip generations" in a way that is mysterious or that gradually phases the trait out. Understanding what is really happening here reinforces why recessive alleles persist.
When a recessive trait appears in one generation, vanishes in the next, and reappears later, it can look as though the trait is fading in and out, perhaps on its way to disappearing. But nothing of the sort is occurring. The allele is present the whole time; it is simply not expressed in the generations where individuals are heterozygous carriers rather than affected homozygotes. A carrier shows the dominant phenotype while carrying and transmitting the recessive allele, so the trait is invisible in that generation but the allele is fully preserved. When two carriers later have a child who is homozygous recessive, the trait reappears, exactly as it was.
This pattern is a normal feature of recessive inheritance, not evidence of decline. The apparent skipping is just the difference between genotype and phenotype playing out across generations: the allele persists continuously in the gene pool while its visible expression comes and goes depending on which genotypes happen to occur. Far from showing a trait disappearing, generation-skipping is a direct demonstration of how recessive alleles endure, hidden in carriers between the occasional appearances of the affected phenotype.
Seeing the Sheltering in Numbers
A quick numerical look makes the carrier-sheltering effect vivid and shows just how thoroughly rare recessive alleles are protected from selection. The comparison is between q², the affected frequency, and 2pq, the carrier frequency, at different allele frequencies.
When the recessive allele frequency q is 0.5, fairly common, q² is 0.25 and 2pq is 0.5, so carriers outnumber affected individuals by only two to one, and a fair share of alleles are exposed in affected homozygotes. But as the allele gets rarer, the picture changes dramatically. At q equal to 0.1, q² is 0.01 and 2pq is about 0.18, so carriers outnumber affected individuals roughly eighteen to one. At q equal to 0.01, q² is 0.0001 and 2pq is about 0.0198, a ratio of nearly two hundred to one. At q equal to 0.001, the ratio approaches two thousand to one.
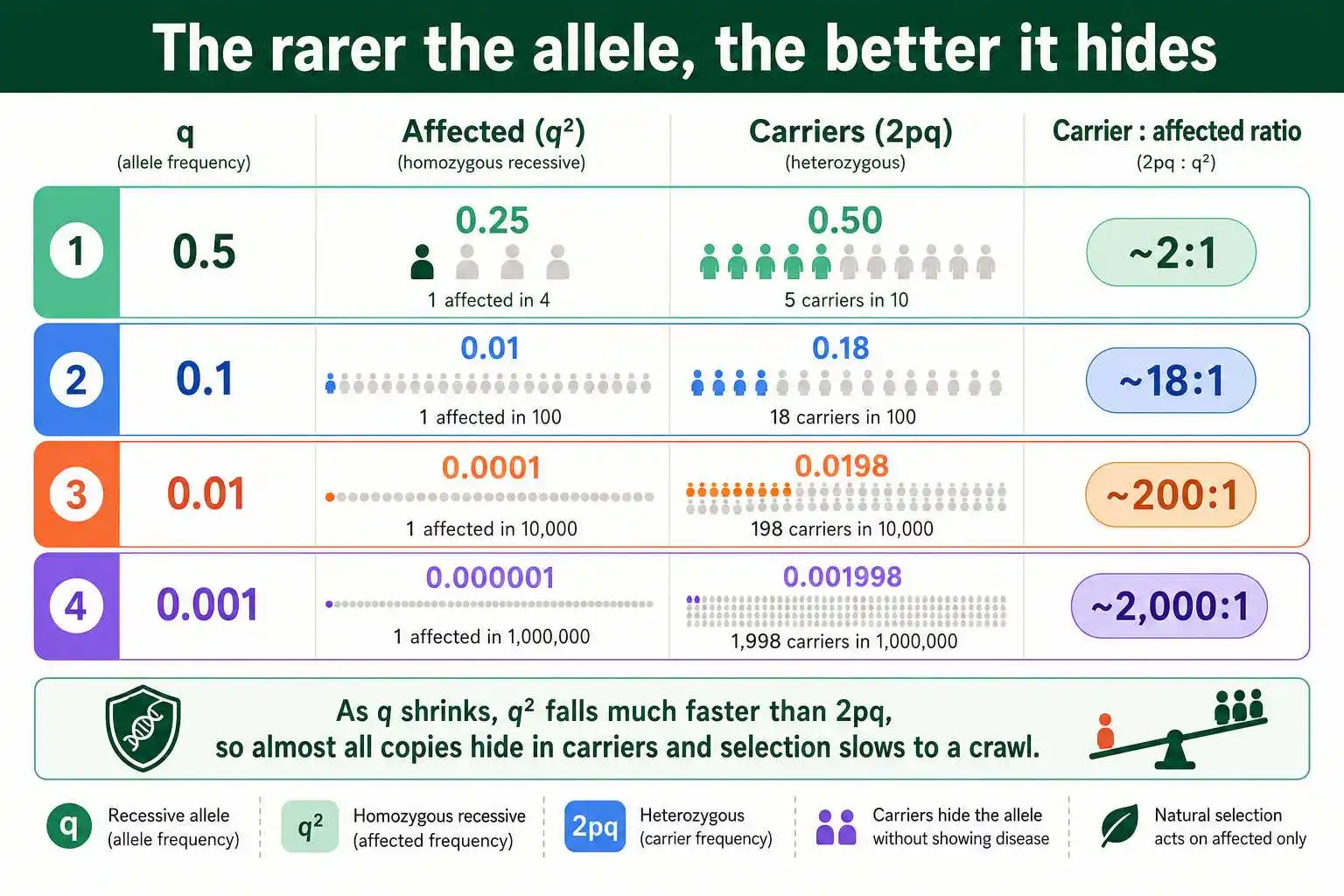
The pattern is unmistakable: the rarer the recessive allele, the more overwhelmingly its copies are concentrated in carriers rather than affected individuals. Since selection can only remove the affected homozygotes, an ever-smaller fraction of the allele's copies are exposed to selection as the allele becomes rare. This is the mathematical heart of why recessive alleles persist. The numbers show that driving a rare recessive allele to extinction through selection is almost impossible, because the allele effectively hides from selection inside the carrier reservoir, and hides better the rarer it gets.
Why This Matters Beyond the Myth
Understanding why recessive traits persist is more than correcting a misconception; it explains real features of genetics and medicine. The persistence of recessive alleles in carriers has direct practical consequences that touch many lives.
The most important is in inherited disease. Because harmful recessive alleles persist sheltered in carriers, recessive genetic disorders continue to appear in populations generation after generation, even though affected individuals may have reduced fitness. This is why carrier screening exists: many healthy people carry recessive disease alleles without any family history, and two carriers can have an affected child. The persistence of these alleles in the carrier reservoir is precisely why such conditions do not simply die out, and why genetic counseling remains relevant across generations.
The persistence of recessive alleles also preserves genetic variation, which has its own importance. A population that retained only dominant alleles would lose a great deal of its genetic diversity, and with it the raw material for future adaptation. By keeping recessive alleles in reserve, mostly hidden in carriers, populations maintain a reservoir of variation that could prove advantageous if conditions change. This conserved variability is one of the deep implications of the Hardy-Weinberg principle, and it connects the persistence of recessive traits to the broader capacity of populations to evolve.
Frequently Asked Questions
Why don't recessive traits disappear over time?
Because dominance describes expression, not frequency, so a dominant allele has no tendency to increase or to eliminate a recessive one. Most copies of a recessive allele hide in healthy carriers where selection cannot act, and under Hardy-Weinberg conditions allele frequencies stay constant across generations.
Does a dominant allele become more common over time?
No. Dominance has no effect on allele frequency. A dominant allele can be rare, like the one causing polydactyly, and a recessive allele can be common. An allele's frequency changes only through evolutionary forces like selection or drift, never because it is dominant.
Why can't selection eliminate a harmful recessive allele?
Because most copies of a rare recessive allele are hidden in heterozygous carriers, invisible to selection, which can only act on affected homozygotes. As the allele gets rarer, even fewer copies are exposed, so selection slows dramatically and the allele persists at low frequency almost indefinitely.
Can recessive alleles ever decrease in frequency?
Yes, but only through evolutionary forces, not because of dominance. Selection against affected individuals, genetic drift in small populations, or gene flow can all change a recessive allele's frequency. The decline, when it happens, is caused by these forces, exactly as it would be for a dominant allele.
The Real Reason They Persist
Recessive traits do not disappear because the intuition behind that expectation is simply wrong. Dominance governs which trait is expressed in a heterozygote, not how common an allele is, so a dominant allele has no inherent tendency to spread and a recessive one has none to vanish. Hardy and Weinberg proved in 1908 that allele frequencies stay constant in the absence of evolutionary forces, making the persistence of recessive alleles the default, not a puzzle.
When selection does act against a recessive trait, the allele still resists elimination because most of its copies hide in healthy carriers, beyond the reach of selection, and the sheltering grows stronger as the allele gets rarer. Recessive alleles change frequency only when a real evolutionary force acts, never from dominance alone, and this persistence keeps disease alleles circulating in carriers while preserving the genetic variation populations need to adapt. You can investigate how carrier and affected frequencies compare for any allele with the Hardy-Weinberg allele frequency calculator. For a clear explanation of why even harmful recessive alleles endure, this resource on recessive alleles from learn-biology is a helpful companion, and the ScienceDirect overview of the Hardy-Weinberg principle details the implication that dominant alleles are not always the most common.